PY17X_0935700 mitogen-activated protein kinase 2, putative (MAP2)
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. yoelii yoelii 17X |
Possible |
RMgm-4816 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-66 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-63 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-62 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
PlasmoGEM (Barseq) | PlasmoGEM | |
| P. berghei ANKA |
Possible |
RMgm-4550 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-4815 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-5343 | Imported from RMgmDB | |
| P. falciparum 3D7 |
Refractory |
22127061 | Theo Sanderson, Wellcome Trust Sanger Institute | |
| P. falciparum 3D7 |
Refractory |
USF piggyBac screen (Insert. mut.) | USF PiggyBac Screen | |
| P. falciparum 3D7 |
Possible |
32681115 With Crispr. Essential for male gametogenesis but not gametocytogenesis. Required to initiate axoneme beating. |
Theo Sanderson, Francis Crick Institute | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. yoelii yoelii 17X | Asexual |
No difference |
RMgm-4816 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Gametocyte |
Difference from wild-type |
RMgm-4816
Normal gametocyte production; no male gamete formation |
Imported from RMgmDB |
| P. berghei ANKA | Asexual |
No difference |
RMgm-66 | Imported from RMgmDB |
| P. berghei ANKA | Asexual |
No difference |
RMgm-63 | Imported from RMgmDB |
| P. berghei ANKA | Asexual |
No difference |
RMgm-62 | Imported from RMgmDB |
| P. berghei ANKA | Asexual |
No difference |
PlasmoGEM (Barseq) | PlasmoGEM |
| P. berghei ANKA | Asexual |
No difference |
RMgm-4815 | Imported from RMgmDB |
| P. falciparum 3D7 | Asexual |
No difference |
32681115 With Crispr. Essential for male gametogenesis but not gametocytogenesis. Required to initiate axoneme beating. |
Theo Sanderson, Francis Crick Institute |
| P. berghei ANKA | Gametocyte |
No difference |
RMgm-63 | Imported from RMgmDB |
| P. berghei ANKA | Gametocyte |
No difference |
RMgm-66 | Imported from RMgmDB |
| P. berghei ANKA | Gametocyte |
No difference |
RMgm-62 | Imported from RMgmDB |
| P. berghei ANKA | Gametocyte |
Difference from wild-type |
RMgm-4815
Normal gametocyte production; no male gamete formation |
Imported from RMgmDB |
| P. falciparum 3D7 | Gametocyte |
Refractory |
32681115 With Crispr. Essential for male gametogenesis but not gametocytogenesis. Required to initiate axoneme beating. |
Theo Sanderson, Francis Crick Institute |
| P. falciparum 3D7 | Gametocyte |
Attenuated |
32681115 (Conditional) | Theo Sanderson, Francis Crick Institute |
| P. berghei ANKA | Ookinete |
Difference from wild-type |
RMgm-63
Male and female gametocyte production is not affected. Genome replication occurs in the male but no nuclear division/ segmentation. No exflagellation of male gametocytes (no male gamete production).No fertilization since male gametocytes do not produce gametes. |
Imported from RMgmDB |
| P. berghei ANKA | Ookinete |
Difference from wild-type |
RMgm-66
Male and female gametocyte production is not affected. No exflagellation observed in mutant clone c2 and drastically reduced in mutant clone b1, therefore, a defect in male gamete formation. |
Imported from RMgmDB |
| P. berghei ANKA | Ookinete |
Difference from wild-type |
RMgm-62
Male and female gametocyte production is not affected. Genome replication occurs in the male but no nuclear division/ segmentation. No exflagellation of male gametocytes (no male gamete production).No fertilization since male gametocytes do not produce gametes. |
Imported from RMgmDB |
Imaging data (from Malaria Metabolic Pathways)
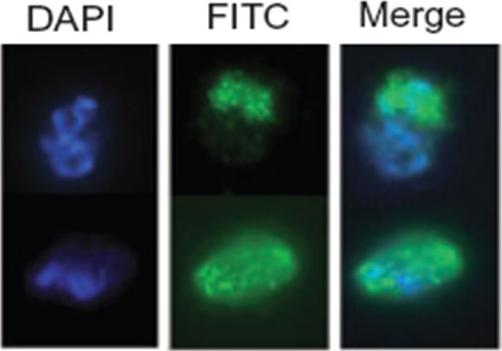
Immunofluorescence. Smears of 3D7 parasites transformed with pCAM-BSD-MAP2HA (containing the 3′ end of the pfmap-2 coding region fused to an haemaglutinin (HA) epitope, followed by the 3′ untranslated region (3′ UTR) from the P. berghei dhfr-ts gene) were subjected to immunofluorescence analysis using an anti-HA antibody.Dorin-Semblat D, Quashie N, Halbert J, Sicard A, Doerig C, Peat E, Ranford-Cartwright L, Doerig C. Functional characterization of both MAP kinases of the human malaria parasite Plasmodium falciparum by reverse genetics. Mol Microbiol. 2007 65:1170-80. Copyright John Wiley & Sons Ltd. 2010.
See original on MMP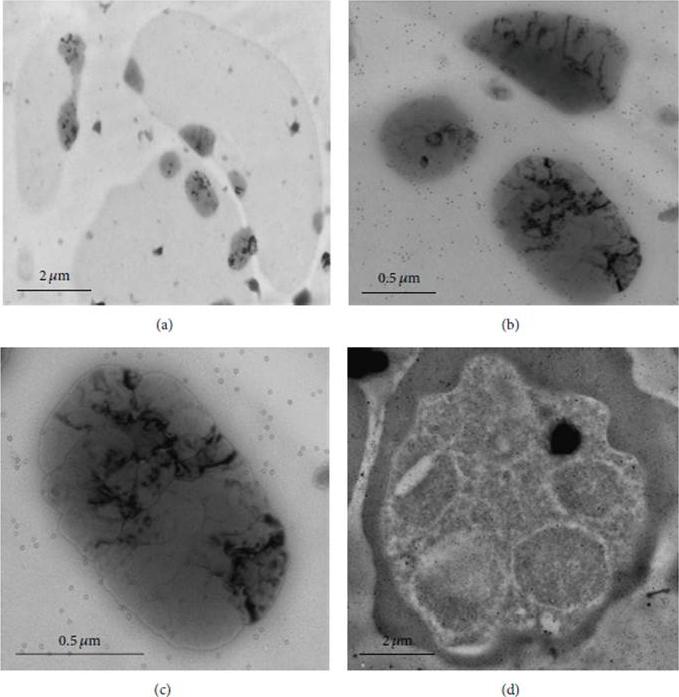
Localization of PfMAP2 by immunoelectron microscopy. Ultrathin sections of P. falciparum parasites at intraerythrocyte stage were labelled with rabbit anti-PPfMAP2 antibody followed with anti-rabbit secondary antibody conjugated with 15 nmcolloidal gold particle. Ultrathin sections of infected erythrocyte were observed under 7 000x (a), 25 000x (b), and 40 000x (c) magnification. Similarly PPfMAP2 staining in the schizont was observed under 8 000x magnification (d). the immunogold was detected in various vesicular structures including nucleus. Since we do not have a specific PPfMAP2 antibody which is nucleus-specific, we have come to understand that the expression of PfMAP2 is regionalised in various organelles of the Plasmodium parasite.Dahalan FA, Sidek HM, Murtey MD, Embi MN, Ibrahim J, Fei Tieng L, Zakaria NA, Abdul-Aziz NM. Phosphorylated and Nonphosphorylated PfMAP2 Are Localized in the Nucleus, Dependent on the Stage of Plasmodium falciparum Asexual Maturation. Biomed Res Int. 2016;2016:1645097.
See original on MMP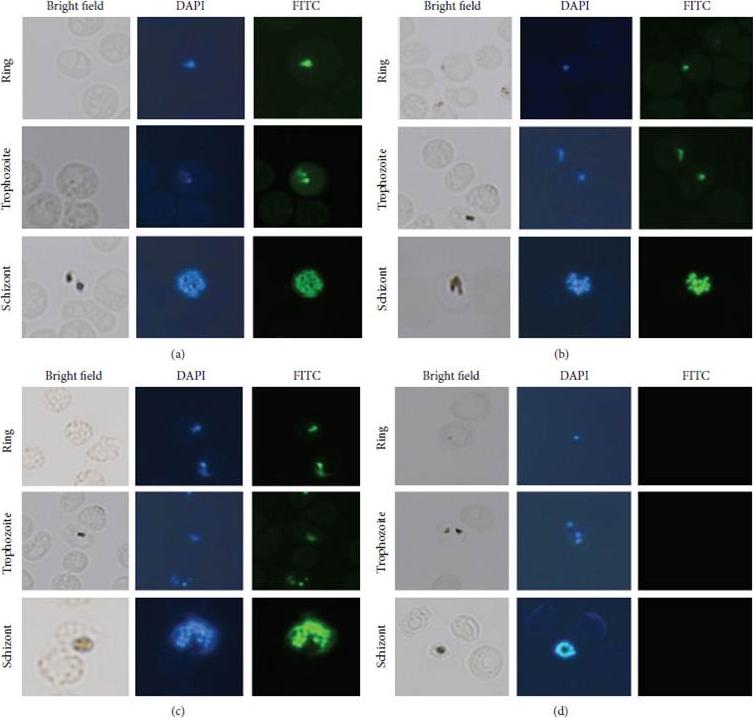
Subcellular distribution of PfMAP2 in P. falciparum. Immunofluorescence analysis of (a) PfMAP2 without phosphatase treatment localization in P. falciparum-infected red blood cells, (b) PfMAP2 with phosphatase treatment localization in P. falciparum-infected red blood cells, (c) PPfMAP2 without phosphatase treatment localization in P. falciparum-infected red blood cells, and (d) PPfMAP2 with phosphatase treatment localization in P. falciparum-infected red blood cells. P. falciparum-infected red blood cells were treated with DAPI (blue) and incubated with primary antibody PfMAP2 (1 : 100) and secondary antibody conjugated-FITC (green) (1 : 200).Dahalan FA, Sidek HM, Murtey MD, Embi MN, Ibrahim J, Fei Tieng L, Zakaria NA, Abdul-Aziz NM. Phosphorylated and Nonphosphorylated PfMAP2 Are Localized in the Nucleus, Dependent on the Stage of Plasmodium falciparum Asexual Maturation. Biomed Res Int. 2016;2016:1645097.
See original on MMPMore information
| PlasmoDB | PY17X_0935700 |
| GeneDB | PY17X_0935700 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | null |
| Orthologs | PBANKA_0933700 , PCHAS_0910600 , PF3D7_1113900 , PKNH_0911500 , PVP01_0914700 , PVX_091340 |
| Google Scholar | Search for all mentions of this gene |