PVX_099930 high molecular weight rhoptry protein 2 (RhopH2)
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. falciparum 3D7 |
Refractory |
21320181 | Theo Sanderson, Wellcome Trust Sanger Institute | |
| P. falciparum 3D7 |
Refractory |
28221136 (CRISPR-mediated) |
Theo Sanderson, Wellcome Trust Sanger Institute | |
| P. falciparum 3D7 |
Refractory |
USF piggyBac screen (Insert. mut.) | USF PiggyBac Screen | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. falciparum 3D7 | Asexual |
Attenuated |
28252383 (Knock down)
Affects new permeability pathways, and hence nutrient availability |
Theo Sanderson, Wellcome Trust Sanger Institute |
| P. falciparum 3D7 | Asexual |
Attenuated |
28221136 (Knock down)
Reduced nutrient uptake |
Theo Sanderson, Wellcome Trust Sanger Institute |
Imaging data (from Malaria Metabolic Pathways)
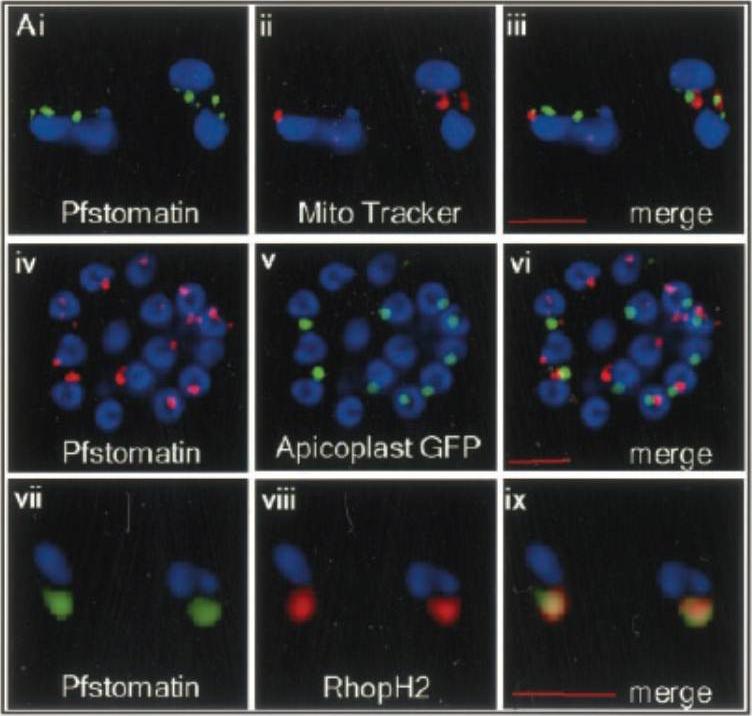
Localization of Pfstomatin to the apical organelles of merozoites and association of parasite rhoptry protein RhopH2 . Indirect immunofluorescence assays of: (i–iii) free merozoites probed with antibodies to Pfstomatin (i; green), and Mito Tracker (ii; red); the relative lack of co-localization of these markers is shown in the merged panel iii. iv–vi intracellular schizont-stage parasites expressing GFP in the apicoplast were probed with antibodies to Pfstomatin (iv; red), GFP (v; green); the relative lack of co-localization of these markers is shown in the merged panel vi. vii–ix free merozoites probed with antibodies to: Pfstomatin (vii; green), RhopH2 (viii; red); the extent of co-localization is shown in the merged panel ix. All images shown are single optical sections. The nucleus (blue) was stained with Hoechst 33342. Scale bars correspond to 2 mm.Hiller NL, Akompong T, Morrow JS, Holder AA, Haldar K. Identification of a stomatin orthologue in vacuoles induced in human erythrocytes by malaria parasites. A role for microbial raft proteins in apicomplexan vacuole biogenesis. J Biol Chem. 2003 278:48413-21.
See original on MMP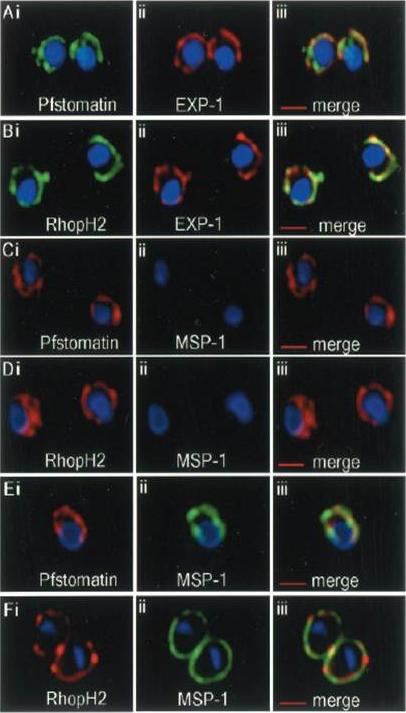
Localization of Pfstomatin and RhopH2 to the PVM in ring-infected erythrocytes (0–8 h post-invasion). Cells were subjected to a standard indirect immunofluorescence assay where samples were permeabilized with saponin and probed with antibodies to: Pfstomatin (Ai; green), RhopH2 (Bi; green), and the PVM marker EXP-1 (Aii and Bii; red); the merged images are shown in Aiii and Biii. In C and D, ring-infected erythrocytes were treated with tetanolysin and incubated with antibodies to: Pfstomatin (Ci; red), RhopH2 (Di; red) and the PPM marker MSP-1 (Cii and Dii; green); the merged images are shown in Ciii and Diii. In E and F, rings treated with tetanolysin and permeabilized with saponin were incubated with antibodies to: Pfstomatin (Ei; red), RhopH2 (Fi; red), and MSP-1 (Eii and Fii; green); the merged images are shown in Diii and Fiii. All images shown are single optical section. The nucleus (blue) was stained with Hoechst 33342. Scale bars represent 2 mm. Hiller NL, Akompong T, Morrow JS, Holder AA, Haldar K. Identification of a stomatin orthologue in vacuoles induced in human erythrocytes by malaria parasites. A role for microbial raft proteins in apicomplexan vacuole biogenesis. J Biol Chem. 2003 278:48413-21.
See original on MMP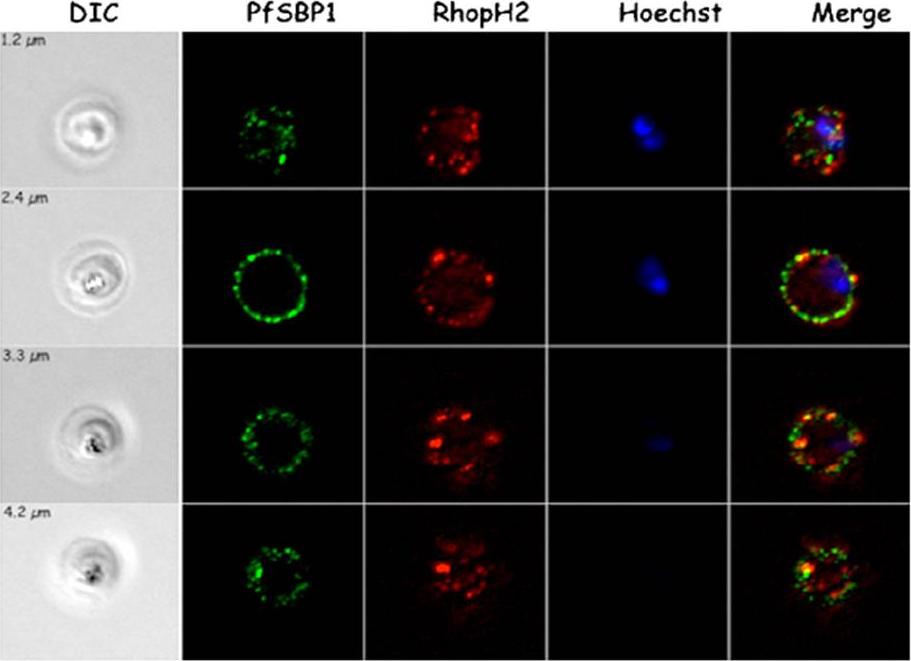
Localization of RhopH2 vs. PfSBP1 in trophozoites. Immunofluorescence studies on SLO-permeabilized trophozoites using RhopH2-specific sera and mouse PfSBP1-specific serum as indicated. The differential interference contrast (DIC) images, the Hoechst nuclei staining and the merge are presented. Four acquisitions at different z-value (as indicated) are presented in each panel. PfSBP1 localizes to the Maurer’s clefts.Vincensini L, Fall G, Berry L, Blisnick T, Braun Breton C. The RhopH complex is transferred to the host cell cytoplasm following red blood cell invasion by Plasmodium falciparum. Mol Biochem Parasitol. 2008 160:81-9. Copyright Elsevier 2009.
See original on MMP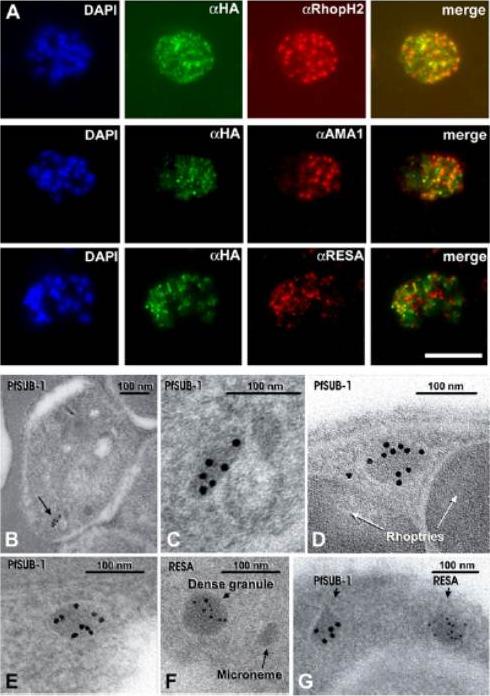
(A) Schizonts of 3D7SUB1HA3 clone C10 dual-labeled with the anti-HA mAb 3F10 (aHA; green) plus mAb 61.3 (aRhopH2), mAb 4G2 (aPfAMA1; micronemes, or mAb 28/2 (aRESA; dense granules. Nuclei were stained with DAPI (blue). Merged images (no DAPI) show that PfSUB1 localizes with none of the other markers. The scale bar represents 5 mm. Identical results were obtained with 3D7SUB1HA3 clone F7 or by using mAbs specific for RAP2 or EBA-175 as markers for rhoptries and micronemes respectively (data not shown). (B–G) Immunoelectron microscopic localization of PfSUB1 in P. falciparum schizonts. (B and C) Labeling of an elongated organelle, arrowed in (B) and at higher magnification in (C), by anti-PfSUB1 antibodies labeled with 10 nm immunogold. (D and E) Two more examples of PfSUB1 positive organelles are shown, also labeled with 10 nm immunogold. These are less elongate than in (B) and (C), reflecting some variation in organelle shape; however, they are typically ellipsoidal, in contrast with the rounded dense granules. (F and G) Staining for RESA with mAb 28/2 and 5 nm immunogold. In (F), an unlabeled microneme is also shown for size comparison. In (G) double staining for PfSUB1 and RESA with different sizes of immunogold shows that the two proteins are in different organelles.Yeoh S, O'Donnell RA, Koussis K, Dluzewski AR, Ansell KH, Osborne SA, Hackett F, Withers-Martinez C, Mitchell GH, Bannister LH, Bryans JS, Kettleborough CA, Blackman MJ. Subcellular discharge of a serine protease mediates release of invasive malaria parasites from host erythrocytes. Cell. 2007 131:1072-83.
See original on MMP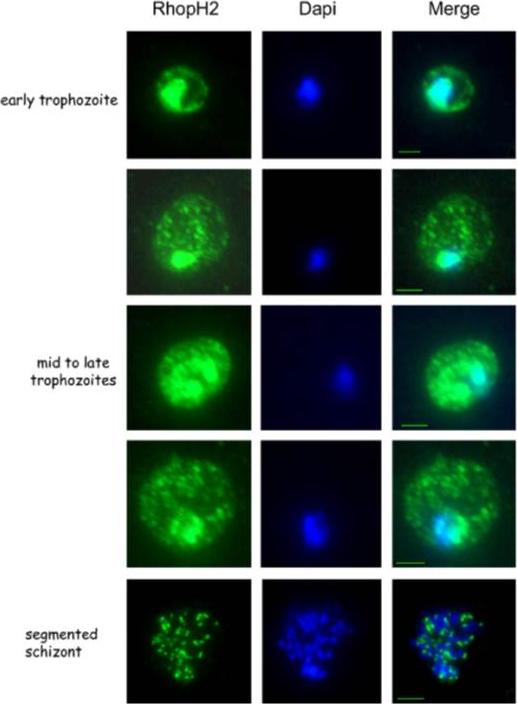
Localisation of RhopH2 at different intra-erythrocytic stages of P. falciparum. Infected erythrocyteswere harvested at different stages of the parasite intra-erythrocytic cycle (early trophozoite, mid to late trophozoites and schizont respectively), acetone/methanol fixed and incubated with the RhopH2-specific antibodies. The nuclei were stained with Dapi (blue). Vincensini L, Fall G, Berry L, Blisnick T, Braun Breton C. The RhopH complex is transferred to the host cell cytoplasm following red blood cell invasion by Plasmodium falciparum. Mol Biochem Parasitol. 2008 160:81-9. Copyright Elsevier 2009.
See original on MMP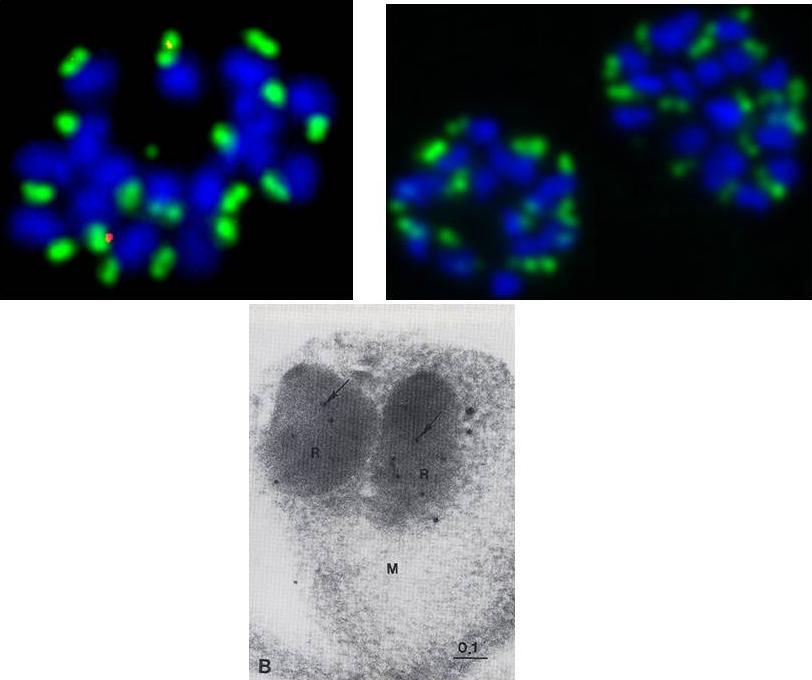
Upper: Localization of RhopH2 in the rhoptries of schizonts with polyclonal and monoclonal Abs.Lower: Electron micrograph of a merozoite (M) reacted with the polyvalent antibodies against the 140 000 mol. wt. protein, followed by Protein A-gold label. Gold particles (arrow) bind specifically to electron dense rhoptries (R) (bar, 0.1 mm).Holder AA, Freeman RR, Uni S, Aikawa M. Isolation of a Plasmodium falciparum rhoptry protein. Mol Biochem Parasitol. 1985 14:293-303.
See original on MMP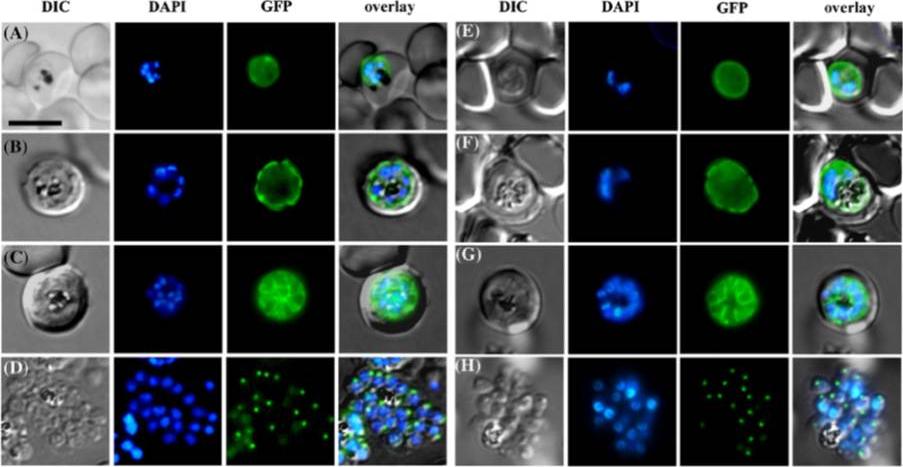
Similar trafficking and subcellular localization of RhopH2-Myc-GFP (A–D) and RhopH2-GFP (E–H) chimeric proteins during late developmental stages of parasite growth. Images were collected from live parasites. (A and E) Early schizont stages showing almost even fluorescence signal in the cytoplasm and probably the parasitophorous vacuole. (B and F) Further developed schizonts showing accumulation of the chimeric proteins around the parasites. (C and G) Later schizont stages showing fluorescence surrounding individual merozoites. Fluorescent vesicles can be seen; probably representing premature rhoptries. (D and H) Segmented schizonts showing concentrated fluorescence signal at the apical end of the formed merozoites. Nuclei were stained with DAPI. Scale bar represents 5 μm.Ghoneim A, Kaneko O, Tsuboi T, Torii M. The Plasmodium falciparum RhopH2 promoter and first 24 amino acids are sufficient to target proteins to the rhoptries. Parasitol Int. 2007 56:31-43. Copyright Elsevier 2009.
See original on MMP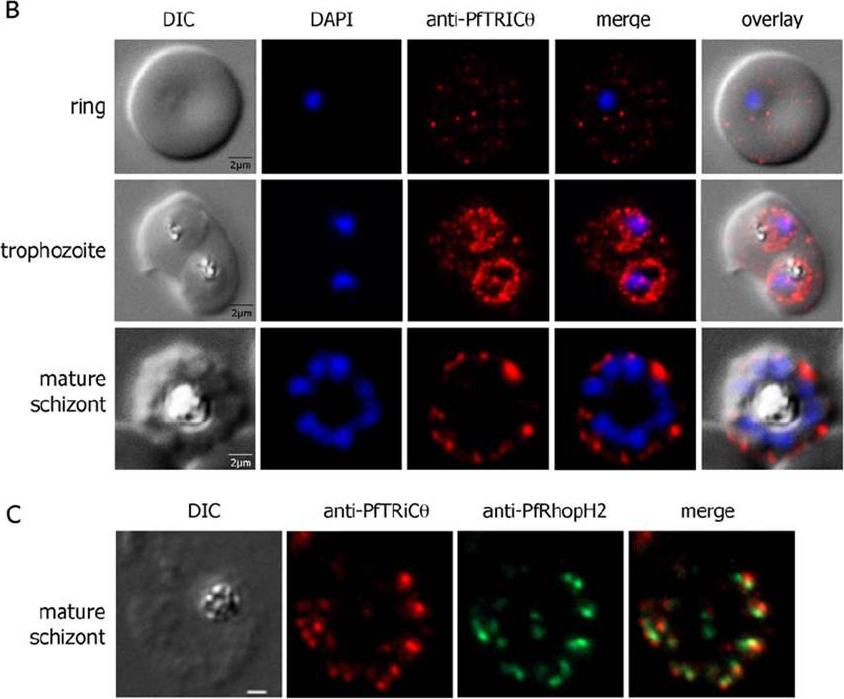
PfTRiC is exported to the cytoplasm of P. falciparum iRBCs. (B). Immunofluorescence studies of erythrocyte infected by Pf3D7 at the early (row 1) and mid (row 2) trophozoite stages and at late schizonte stage using PfTRiCθ specific antibodies (in red). DAPI: DAPI staining of the parasite nucleus (in blue). (C) Double IFA labelling of PfTRiCq (in red) and the rhoptry protein PfRhopH2 (in green) in segmented schizonts using specific sera and methanol-acetone fixed infected erythrocytes .Mbengue A, Vialla E, Berry L, Fall G, Audiger N, Demettre-Verceil E, Boteller D, Braun-Breton C. New export pathway in Plasmodium falciparum-infected erythrocytes: role of the parasite group II chaperonin, PfTRiC. Traffic. 2015 Jan 23. [Epub ahead of print]
See original on MMP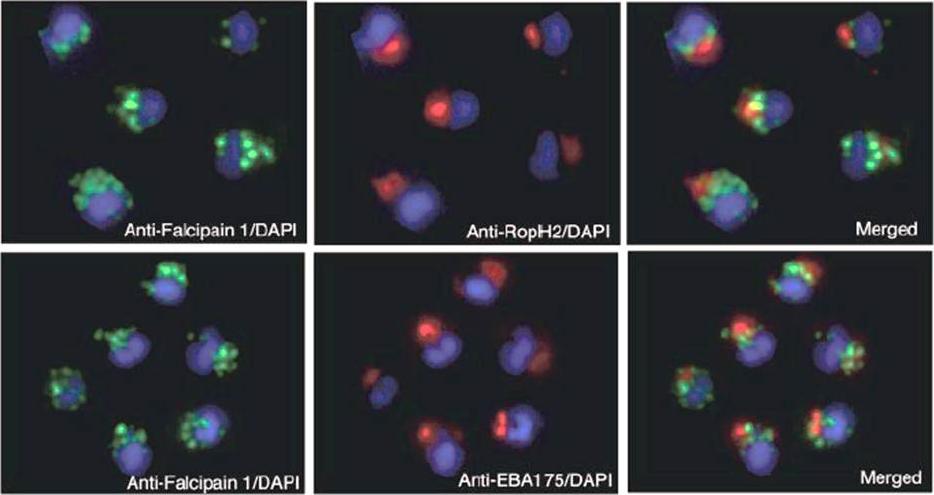
Localization of falcipain 1 (FP1) in merozoites by immunofluorescence imaging. Samples of merozoites were smeared and fixed in 1% paraformaldehyde. The slides were incubated with an antibody to falcipain 1 or with antibodies specific to the rhoptry (RopH2) and microneme (EBA-175) proteins. Nuclei were stained with 4’,6-diamidino-2-phenylindole (DAPI). The lack of colocalization indicates that falcipain 1 is found in compartments at the apical end of the merozoite that are distinct from the rhoptries and micronemes.Greenbaum DC, Baruch A, Grainger M, Bozdech Z, Medzihradszky KF, Engel J, DeRisi J, Holder AA, Bogyo M. A role for the protease falcipain 1 in host cell invasion by the human malaria parasite. Science. 2002 298(5600):2002-6.
See original on MMP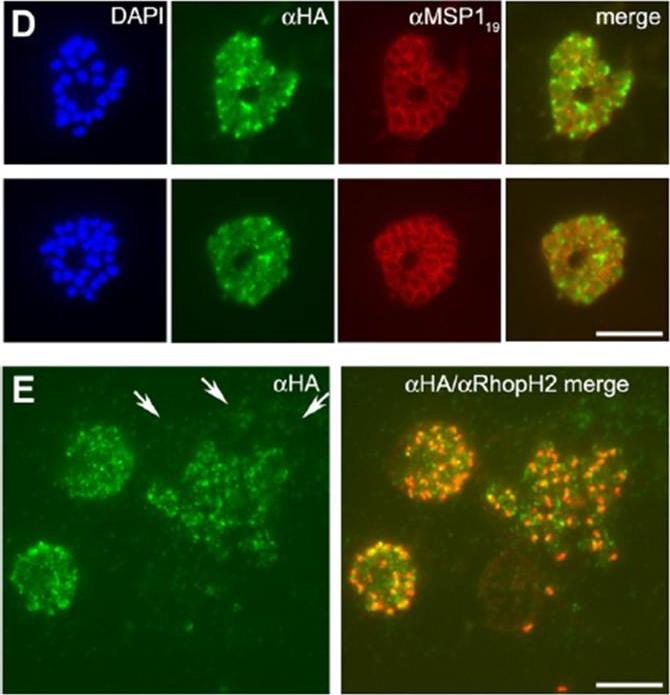
D) Segmented schizonts of 3D7SUB1HA3clone C10 dual-labeled with mAb 3F10 (aHA; green) and mAb 1E1 (aMSP119; red). Merged images (no DAPI) show partial colocalization of PfSUB1HA3 and MSP1. Dual labeling suggestisthat near the end of schizogony PfSUB1 can translocate from its previous organellar location into the PV. (E) A ‘‘cloud’’ of PfSUB1HA3 around a bursting schizont of 3D7SUB1HA3 clone C10 dual-labeled with mAb 3F10 (aHA; green) and mAb 61.3 (aRhopH2). The right-hand panel shows the merged image. The scale bar represents 5 mm. Where bursting schizonts were visible, the free merozoites were surrounded by a ‘‘cloud’’ of anti-HA signal, suggesting that at the point of egress PfSUB1 discharge had already begun.Yeoh S, O'Donnell RA, Koussis K, Dluzewski AR, Ansell KH, Osborne SA, Hackett F, Withers-Martinez C, Mitchell GH, Bannister LH, Bryans JS, Kettleborough CA, Blackman MJ. Subcellular discharge of a serine protease mediates release of invasive malaria parasites from host erythrocytes. Cell. 2007 131:1072-83.
See original on MMP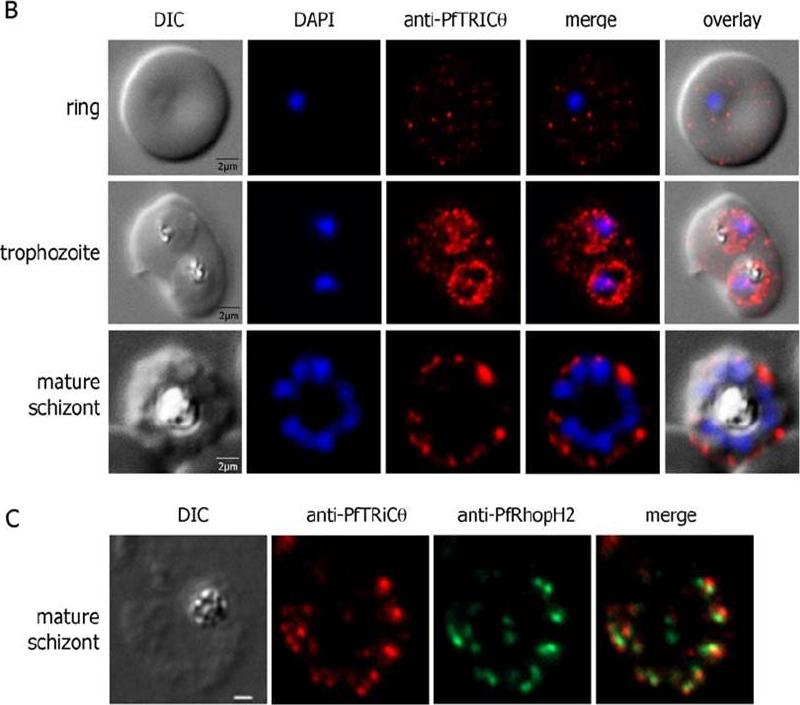
PfTRiC is exported to the cytoplasm of P. falciparum iRBCs. (B) Immuno-fluorescence studies of erythrocyte infected by Pf3D7 at the early (row 1) and mid (row 2) trophozoite stages and at late schizonte stage using PfTRiCθ specific antibodies (in red). DAPI: DAPI staining of the parasite nucleus (in blue). (C) Double IFA labelling of PPfTRiCθ (in red) and the rhoptry protein PfRhopH2 (in green) in segmented schizonts using specific sera and methanol-acetone fixed infected erythrocytes. PfTRiCθ was detected in the iRBC cytosol from early rings to early schizonts. Labelling of the parasite (and possibly the parasitophorous vacuole) was instead observed for mature stages (mature trophozoites to schizonts). Moreover, following the individualization of merozoites, PfTRiCθ was only detected at the apex of each merozoite. Double labelling immunofluorescence experiments using anti-PfTRiCθ and anti-RhopH2 antibodies.Mbengue A, Vialla E, Berry L, Fall G, Audiger N, Demettre-Verceil E, Boteller D, Braun-Breton C. New Export Pathway in Plasmodium falciparum-Infected Erythrocytes: Role of the Parasite Group II Chaperonin, PfTRiC. Traffic. 201516(5):461-75.
See original on MMP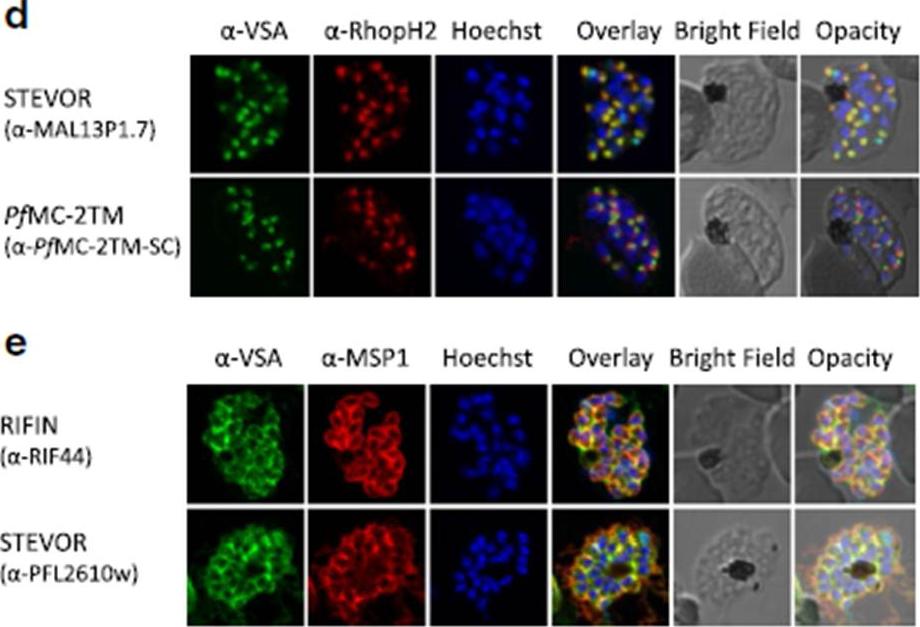
d Co-localization of STEVOR α-MAL13P1.7 or α-PfMC-2TM-SC (green) with the rhoptry marker RhopH2 (red). e Co-localization of α-RIF44 and STEVOR α-PFL2610w (green) with the merozoite surface protein MSP1 (red). Variant surface antigens (VSA).Bachmann A, Scholz JA, Janßen M, Klinkert MQ, Tannich E, Bruchhaus I, Petter M. A comparative study of the localization and membrane topology of members of the RIFIN, STEVOR and PfMC-2TM protein families in Plasmodium falciparum-infected erythrocytes. Malar J. 2015 Jul 14:274.
See original on MMP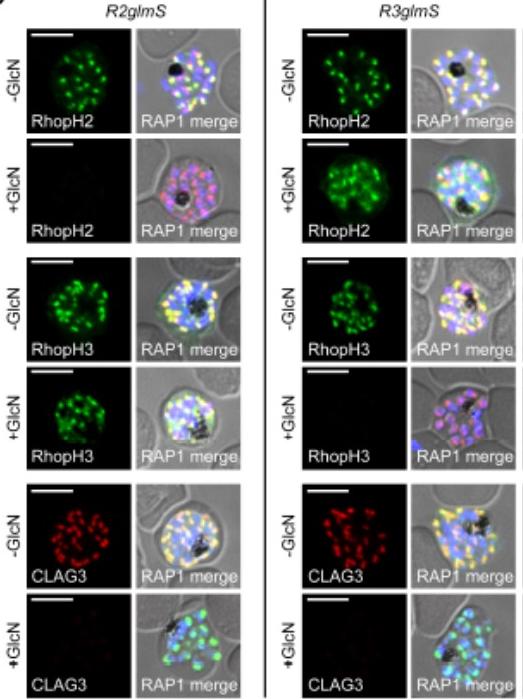
IFA of indicated proteins in R2glmS (RopH2 knockdown) and R3glmS (RopH3 knockdown) at schizont stage; colocalization with RAP1 and apical puncta in daughter merozoites indicates rhoptry trafficking. Scale bars, 5 μm. GlcN treatment of either R2glmS or R3glmS abolished not only the targeted gene product but also CLAG3, a third better-characterized member of the RhopH complex. Indirect immunofluorescence microscopy confirmed this surprising finding and provided additional insights. While RhopH2 was still detected in the R3glmS knockdown, it did not fully colocalize with RAP1 but instead had a more diffuse distribution; this suggests RhopH2 may be retained in the parasite endoplasmic reticulum if it fails to interact with RhopH3. In contrast, RhopH3 appeared to traffic normally inGlcN-treated R2glmS parasites, as indicated by apical colocalization with RAP1 inmerozoites. Thus, CLAG3, RhopH2 and RhopH3 assemble into a complex either during or immediately after translation.Ito D, Schureck MA, Desai SA. An essential dual-function complex mediates erythrocyte invasion and channel-mediated nutrient uptake in malaria parasites. Elife. 2017 Feb 21;6.
See original on MMP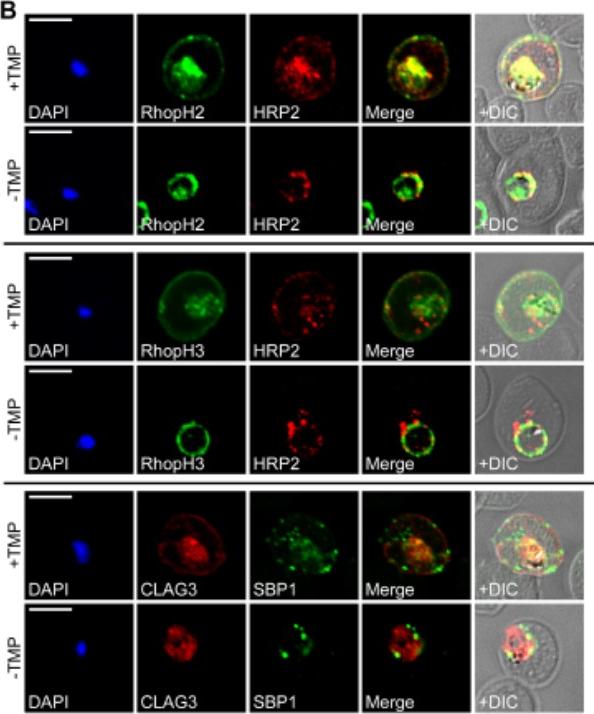
RhopH proteins are deposited in the PV and exported via PTEX. IFA of 13F10 showing export of RhopH proteins after growth with or without TMP for 48 h. Colocalization with exported HRP2 or SBP1, as indicated. Scale bars, 5 μm. Export of RhopH2 and RhopH3 were both fully blocked in trophozoite-stage parasites grown for 48 h without TMP. Colocalization with HRP2, a parasite protein carrying the canonical PEXEL export signal showed both proteins trapped in the vacuole upon PTEX knockdown. CLAG3 export requires PTEX activity as TMP removal prevented its movement out of the PV (bottom row); colocalization with SBP1, another PTEX substrate.Ito D, Schureck MA, Desai SA. An essential dual-function complex mediates erythrocyte invasion and channel-mediated nutrient uptake in malaria parasites. Elife. 2017 Feb 21;6.
See original on MMP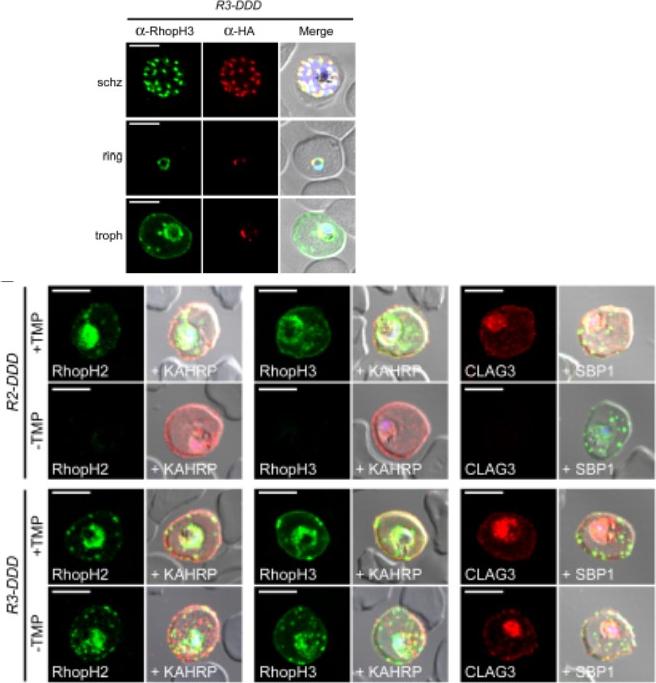
(D) IFA of R3-DDD (conditional RopH3 mutant) probed with indicated antibodies at each parasite stage. Note that both antibodies detect protein in schizont rhoptries, but that only the anti-RhopH antibody detects export to the host membrane in trophozoites. Scale bars, 5 μm. (E) Trophozoite-stage IFAs of indicated parasites cultivated with and without TMP for 48 h, showing that knockdown abolishes each member of the complex in R2-DDD (conditional RopH2 mutant) but not R3-DDD. Co-localization is shown with exported parasite proteins KAHRP or SBP1 (red or green in merge panels, respectively). Scale bars, 5 μm. The RhopH complex appears to be secreted into the PV (upper panel), suggesting that its member proteins must cross the bounding PVM to enter host cytosol and eventually reach the erythrocyte surface. While TMP removal rendered all three RhopH proteins undetectable in the R2-DDD parasite, these proteins and their transfer to trophozoites were preserved inR3-DDD.Ito D, Schureck MA, Desai SA. An essential dual-function complex mediates erythrocyte invasion and channel-mediated nutrient uptake in malaria parasites. Elife. 2017 Feb 21;6. PMID: 28221136.
See original on MMP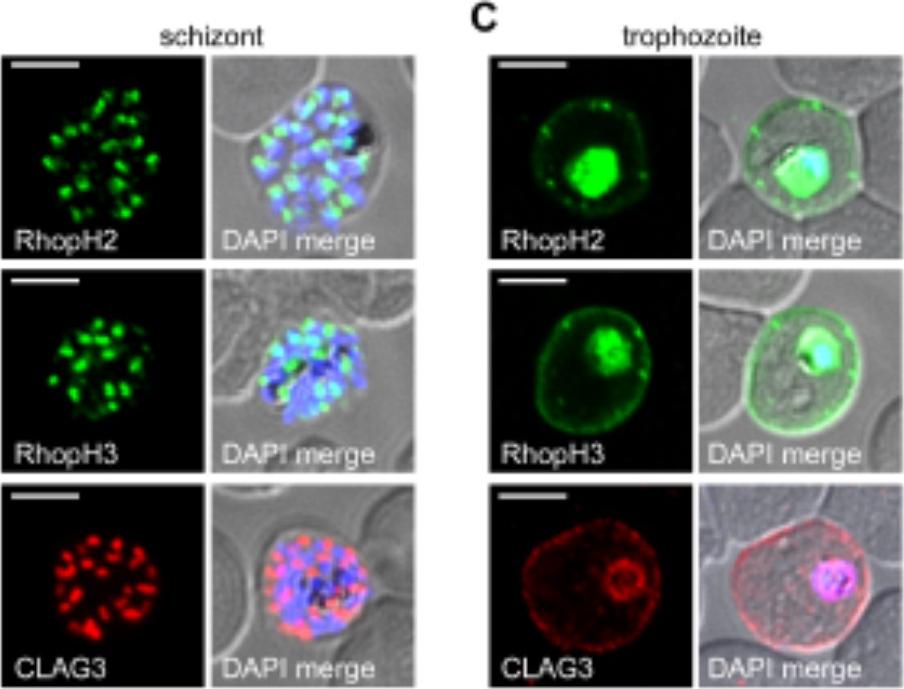
RhopH2 and RhopH3 are membrane-associated proteins. IFA of indicated proteins in R2glmS (RopH2 knockdown) and R3glmS (RopH3 knockdown) at the trophozoite stage. IFA of mature schizont-infected cells with indicated antibodies. Punctate labeling indicates that these proteins localize to rhoptry organelles in daughter merozoites. Scale bar, Scale bars, 5 μm. IFA of trophozoite-infected cells, showing that each protein localizes to the host membrane and to the vacuole surrounding the intracellular parasite. Scale bar, 5 μmImmunofluorescence microscopy confirmed packaging into rhoptries within schizont-infected cells (B). Imaging of maturing trophozoite-stage parasites also showed that these proteins traffic to the host membrane (C). Because each gene is transcribed and translated only within schizonts, the protein found in these maturing trophozoites reflects transfer from the merozoite during invasion.Ito D, Schureck MA, Desai SA. An essential dual-function complex mediates erythrocyte invasion and channel-mediated nutrient uptake in malaria parasites. Elife. 2017 Feb 21;6.
See original on MMP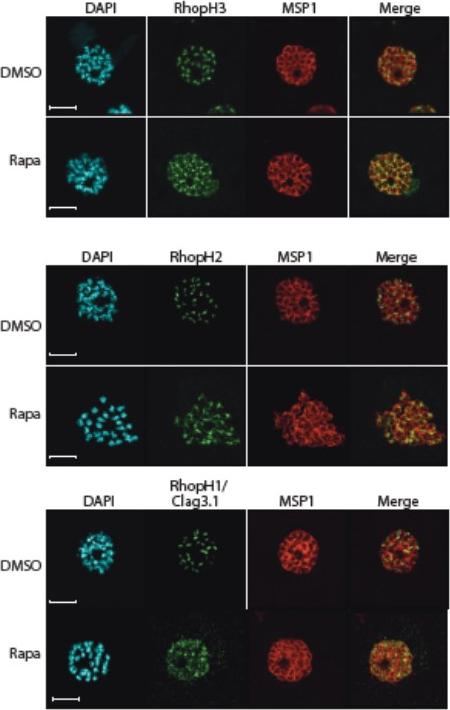
Truncation of RhopH3 leads to mistrafficking of components of the RhopH complex. IFA of mature schizonts of control (DMSO) and rapamycin-treated rhopH3-loxP parasites, probed with MSP1-specific antibodies (either mAb 89.1 or rabbit polyclonal anti-MSP1 antibodies; red) and antibodies to the three indicated RhopH components (green). Mis-localisation of the RhopH proteins was observed in all cases, and in the case of RhopH3 the protein often appeared to reside external to the plasma membrane of intracellular merozoites. Parasite nuclei were visualized by staining with DAPI. Note that, for clarity, the merge panels do not include the DAPI signal. Scale bar, 5 μm.Sherling ES, Knuepfer E, Brzostowski JA, Miller LH, Blackman MJ, van Ooij C. The Plasmodium falciparum rhoptry protein RhopH3 plays essential roles in host cell invasion and nutrient uptake. Elife. 2017 6. pii: e23239.
See original on MMP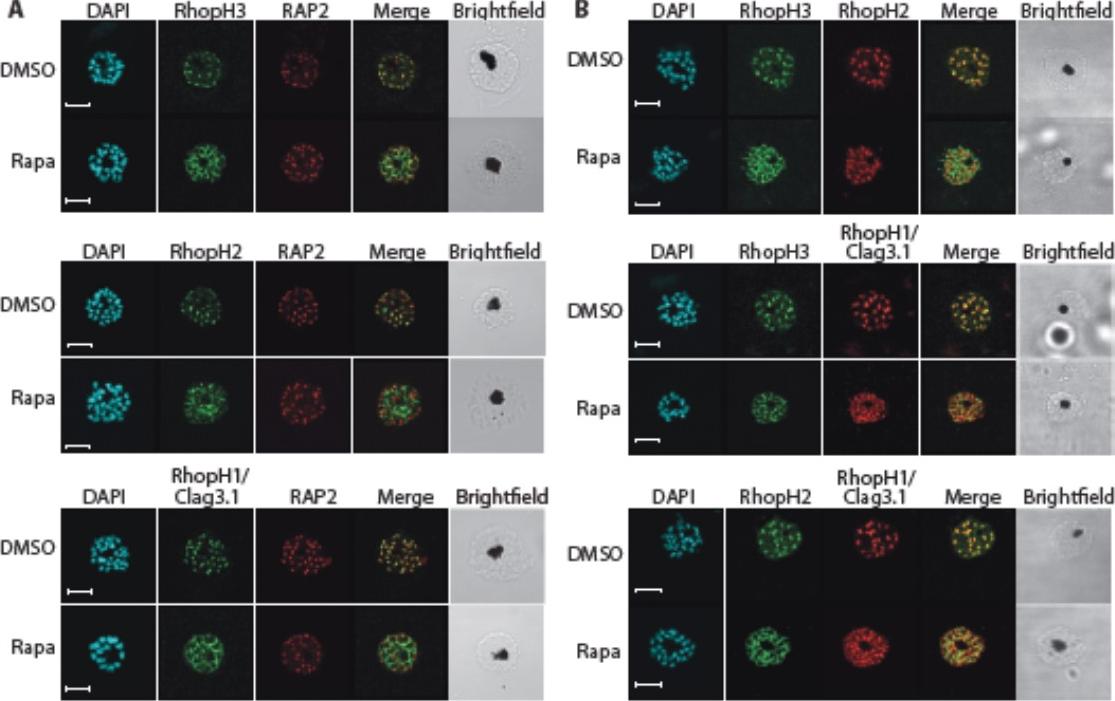
Truncation of RhopH3 leads to mistrafficking of components of the RhopH complex and loss of complex formation. A) IFA showing colocalization of RhopH3, RhopH2 and RhopH1/Clag3.1 with the rhoptry marker RAP2 in schizonts of control (DMSO) rhopH3-loxP parasites but loss of colocalization following rapamycin (Rapa) treatment. Parasite nuclei were visualized by staining with 4,6-diamidino-2-phenylindole (DAPI). Scale bar, 5 μm. Immunofluorescence analysis (IFA) showed that, as expected, RhopH3 colocalized with the rhoptry marker RAP2 in mature schizonts of control rhopH3-loxP parasites. However, in rapamycin-treated (RhopH3Δ4-219 6) parasites, this colocalization was lost, although RAP2 was still detected in a punctate, apically-disposed pattern typical of rhoptries B) Colocalization of the members of the RhopH complex. RhopH3, RhopH2 and Rhop 1/Clag3.1 colocalize in rhopH3-loxP parasites treated with DMSO, but this colocalization is lost in parasites treated with rapamycin. Scale bar, 2 μm. Neither RhopH2 nor RhopH1/Clag3.1 colocalized with 233 RhopH3Δ4-6 in the mutant parasites. The RhopH2 and RhopH1/Clag3.1 signals were also distinct in the mutant parasites, although in this case some limited colocalization of these proteins was apparent (bottom images).Sherling ES, Knuepfer E, Brzostowski JA, Miller LH, Blackman MJ, van Ooij C. The Plasmodium falciparum rhoptry protein RhopH3 plays essential roles in host cell invasion and nutrient uptake. Elife. 2017 6. pii: e23239.
See original on MMP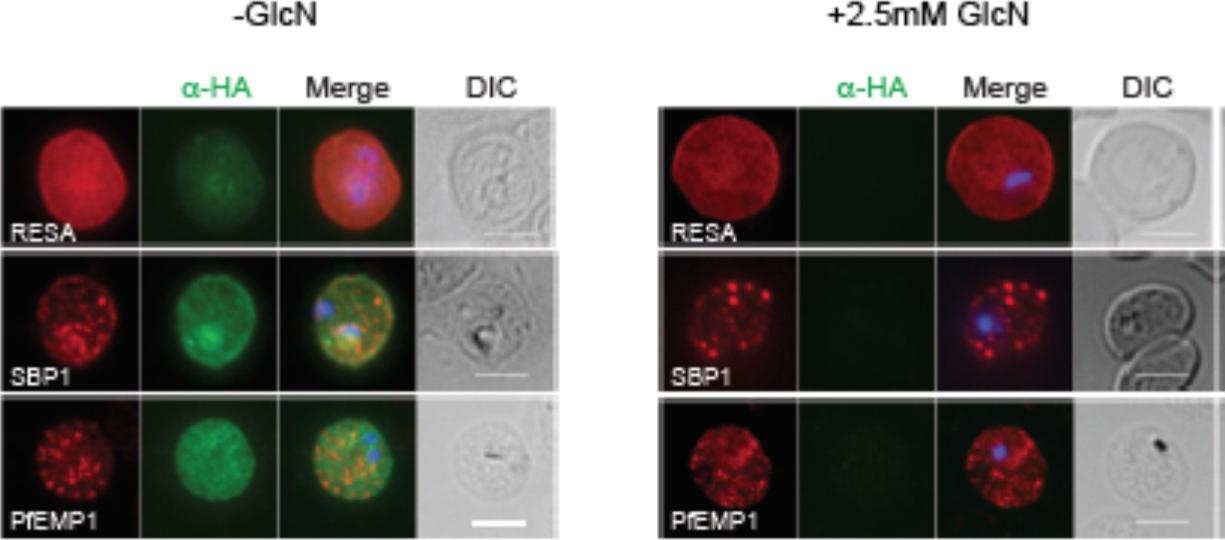
RhopH2 is not involved in the trafficking of exported proteins in the erythrocyte cytoplasm. (a) Representative IFAs of erythrocytes infected with RhopH2-HAglmS parasites grown in 0 mM or 2.5 mM GlcN using the indicated antibodies show trafficking of RESA, SBP1 and PfEMP1 is unaffected upon RhopH2 knockdown. Scale bar = 5 μm. No defect in the export of PfEMP1, or trafficking of either RESA to the erythrocyte membrane or SBP1 to the Maurer’s clefts was evident after knocking down RhopH2 expression with GlcN.Counihan N, Chisholm SA, Bullen HE, Srivastava A, Sanders PR, Jonsdottir TK, Weiss GE, Ghosh S, Crabb BS, Creek DJ, Gilson PR, de Koning-Ward TF. Plasmodium falciparum parasites deploy RhopH2 into the host erythrocyte to obtain nutrients, grow and replicate. Elife. 2017 Mar 2;6. pii: e23217.
See original on MMP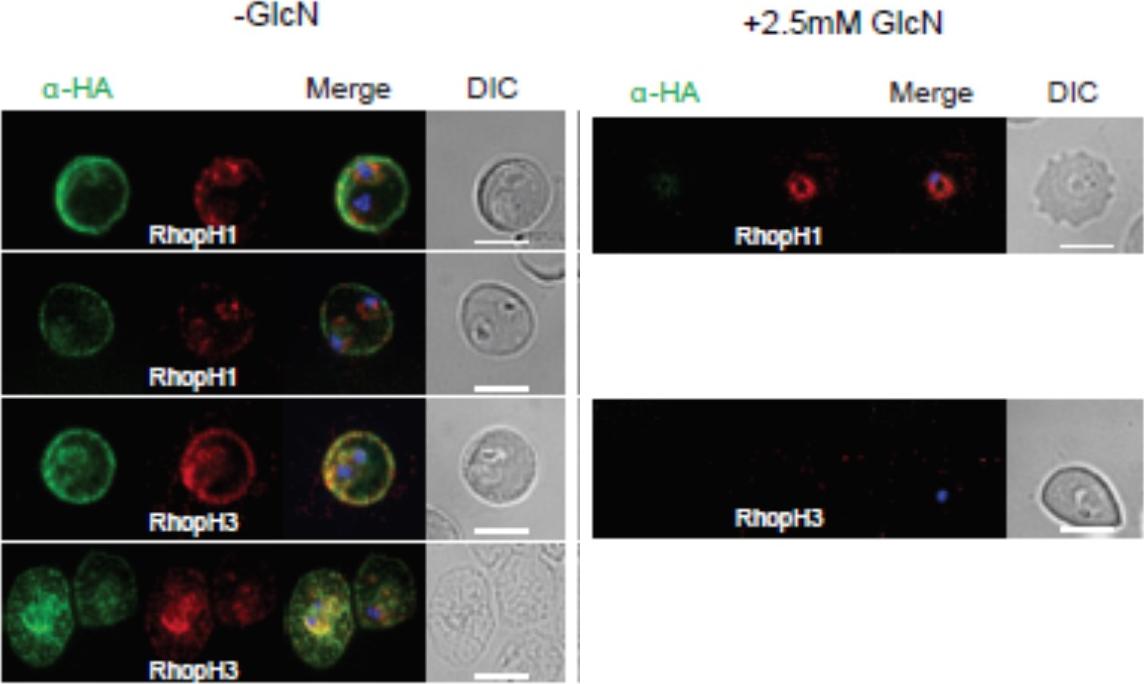
Localization of RhopH1/clag3 and RhopH3 in infected erythrocytes when RhopH2 expression is knocked down. Representative immunofluorescence analysis of erythrocytes infected with RhopH2-HAglmS parasites grown in 0 mM or 2.5 mM GlcN. Cells fixed with acetone/methanol and labelled with anti-HA antibody to detect RhopH2 IFAs and other antibodies, as indicated. the localization of RhopH3 and to a lesser extent RhopH1/clag3 was affected when RhopH2 expression was knocked down.Counihan N, Chisholm SA, Bullen HE, Srivastava A, Sanders PR, Jonsdottir TK, Weiss GE, Ghosh S, Crabb BS, Creek DJ, Gilson PR, de Koning-Ward TF. Plasmodium falciparum parasites deploy RhopH2 into the host erythrocyte to obtain nutrients, grow and replicate. Elife. 2017 Mar 2;6. pii: e23217
See original on MMP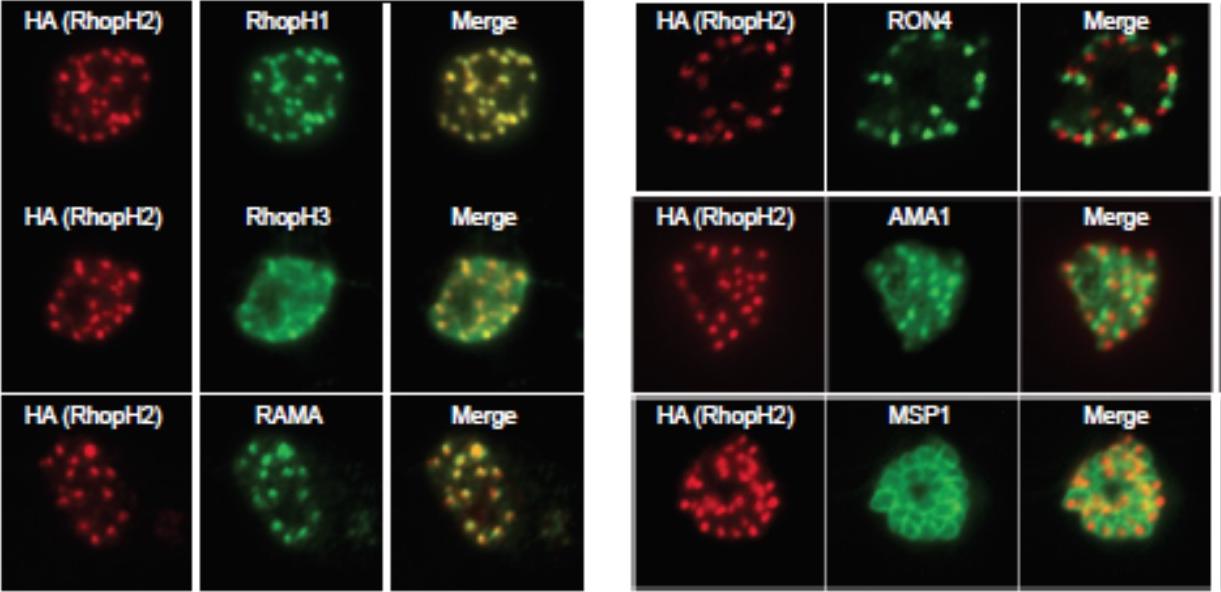
Generation of transgenic parasites in which RhopH2 is epitope-tagged. Immunofluorescence analysis (IFA) on schizonts fixed with acetone/methanol and labelled with anti-HA antibody to detect RhopH2 and other antibodies, as indicated. Immunofluorescence analysis (IFA) confirmed RhopH2-HA localized to the rhoptry and co-localized with other rhoptry bulb proteins, RhopH1, RhopH3 and RAMA but not with the rhoptry neck protein, RON4, the micronemal marker, AMA-1 or the plasma membrane protein MSP1.Counihan N, Chisholm SA, Bullen HE, Srivastava A, Sanders PR, Jonsdottir TK, Weiss GE, Ghosh S, Crabb BS, Creek DJ, Gilson PR, de Koning-Ward TF. Plasmodium falciparum parasites deploy RhopH2 into the host erythrocyte to obtain nutrients, grow and replicate. Elife. 2017 Mar 2;6. pii: e23217
See original on MMP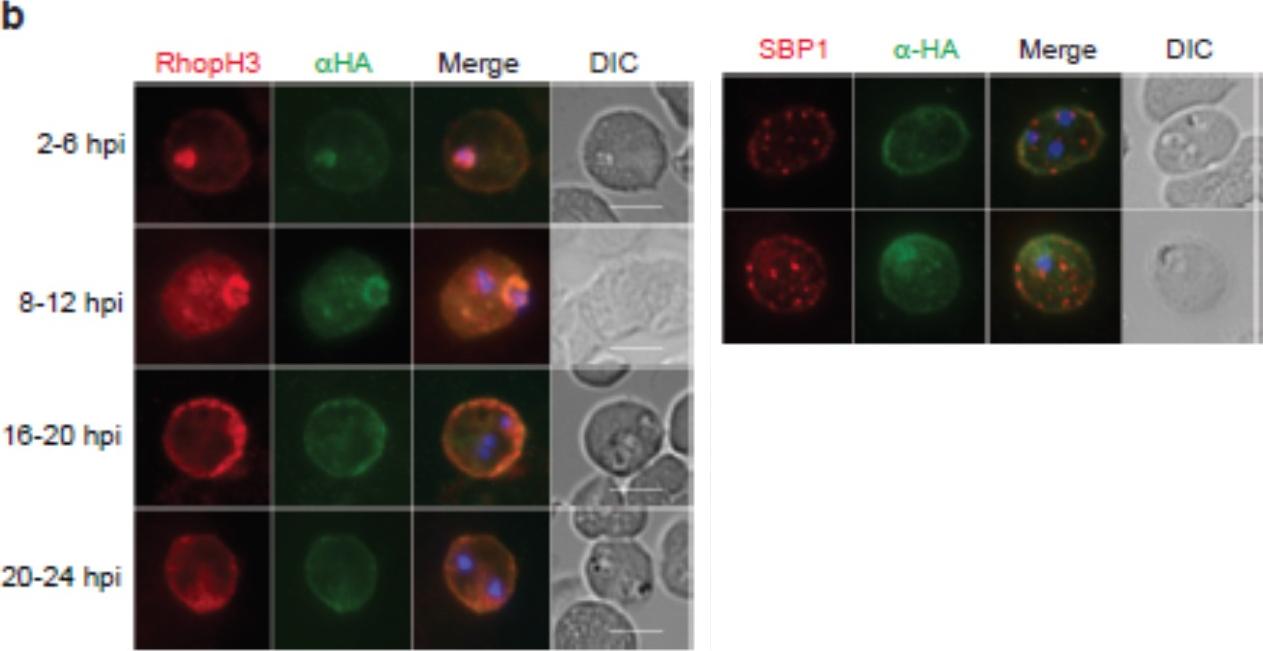
Expression, localisation and solubility profile of P. falciparum RhopH2. Immunofluorescence analysis (IFA) on erythrocytes infected with PfRhopH2-HAglmS and fixed with acetone/methanol. RhopH2 is labeled with the anti-HA antibody. The bars represent 5 μm. (c) IFA on erythrocytes infected with PfRhopH2-HAglmS, fixed with acetone/methanol and probed with anti-HA (for RhopH2) and antibodies to the Maurer’s cleft protein SBP1 show that RhopH2 and SBP1 do not co-localize.Counihan N, Chisholm SA, Bullen HE, Srivastava A, Sanders PR, Jonsdottir TK, Weiss GE, Ghosh S, Crabb BS, Creek DJ, Gilson PR, de Koning-Ward TF. Plasmodium falciparum parasites deploy RhopH2 into the host erythrocyte to obtain nutrients, grow and replicate. Elife. 2017 Mar 2;6. pii: e23217.
See original on MMP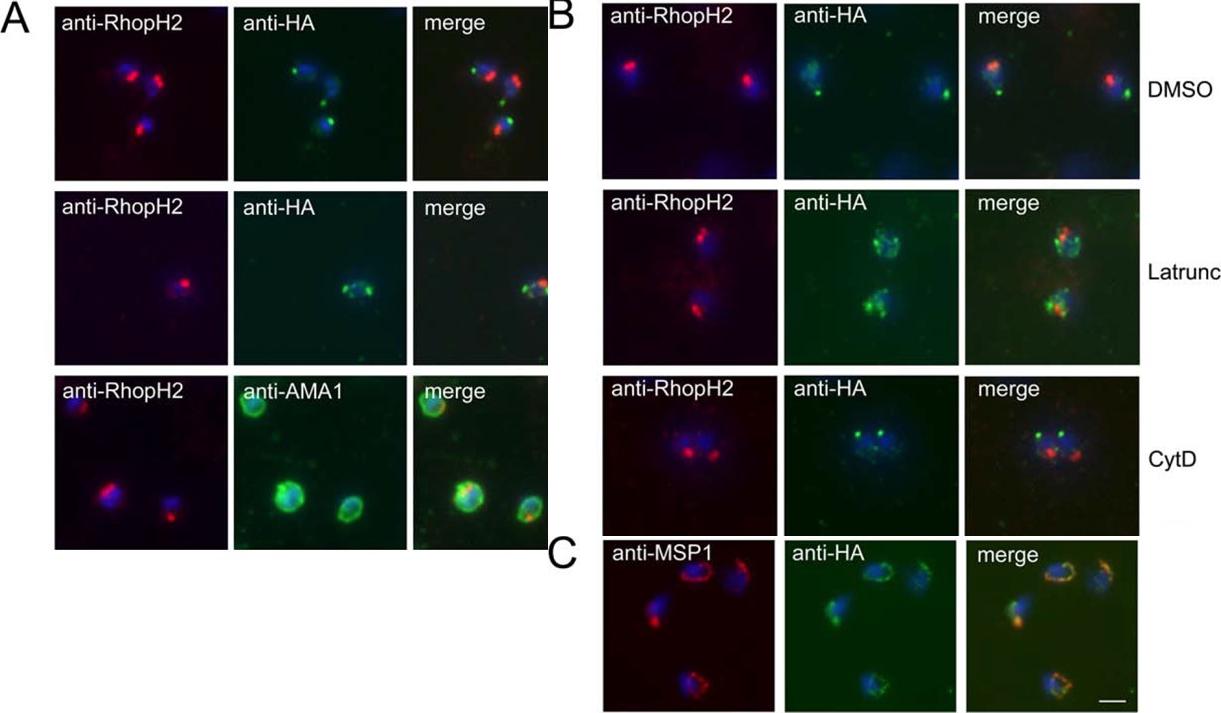
PfSUB2 Redistributes to the Free Merozoite Posterior in an Actin-Dependent Manner. (A) IFA images of free merozoites of PfSUB2HA clone 2D dual-labelled with mAb 61.3 (anti-RhopH2) and either mAb 3F10 (anti-HA) or 4G2 (anti-AMA1). The anti-HA signal formed either a single punctate signal at the extreme posterior of the merozoite (top), or twin foci lateral to its anterior–posterior axis (middle). Identical results were obtained using PfSUB2HA clone 10E, or using anti-RAP2 mAb H5 as the rhoptry marker (not shown). (B) Latrunculin inhibits rearward translocation of PfSUB2. Merozoites of PfSUB2HA clone 2D released into medium containing 1% (v/v) DMSO only (solvent control), or 5 mM latrunculin A (Latrunc), or 4 mM cytochalasin D (Cyt D), probed as above with mAb 61.3 (anti-RhopH2) and mAb 3F10 (anti-HA). Note that neither compound had any effect on the efficiency of schizont rupture and merozoite release, nor on AMA1 relocalisation, but both blocked erythrocyte invasion by more than 95%at the concentrations used (not shown). (C) PfSUB2 remains at the plasma membrane of the newly invaded parasite. Ring stages of PfSUB2HA clone 2D (<2 h post-invasion) probed with anti-MSP1 mAb 1E1 and anti-HA mAb 3F10. Parasite nuclei are stained throughout with DAPI (blue). Scale bar represents 2 mm.Harris PK, Yeoh S, Dluzewski AR, O'Donnell RA, Withers-Martinez C, Hackett F, Bannister LH, Mitchell GH, Blackman MJ. Molecular identification of a malaria merozoite surface sheddase. PLoS Pathog. 2005 Nov;1(3):241-51.
See original on MMP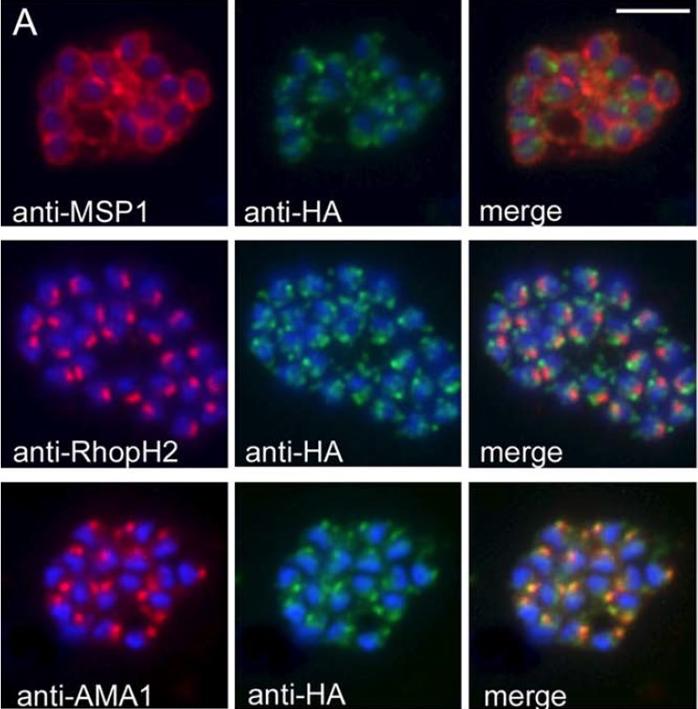
PfSUB2 Is a Microneme Protein. (A) IFA images of schizonts of PfSUB2HA clone 2D dual-labelled with mAbs X509 (anti-MSP1), 61.3 (anti-RhopH2), or 4G2 (anti-AMA1), plus in each case mAb 3F10 (anti-HA) The anti-HA signal co-localised only with the anti-AMA1 signal. Identical results were obtained with the unclonedtransgenic PfSUB2HA line, and/or when anti-RAP2 mAb H5 was used as the rhoptry marker instead of mAb 61.3 (not shown). Parasite nuclei are stained throughout with DAPI (blue). Scale bar represents 2 mm. the anti-HA signal was quite different from that of MSP1, it was adjacent to but distinct from that of the rhoptry specific signals, but it co-localised completely with that ofAMA1, indicating that in mature schizonts, PfSUB2 accumulates in micronemes.Harris PK, Yeoh S, Dluzewski AR, O'Donnell RA, Withers-Martinez C, Hackett F, Bannister LH, Mitchell GH, Blackman MJ. Molecular identification of a malaria merozoite surface sheddase. PLoS Pathog. 2005 Nov;1(3):241-51.
See original on MMP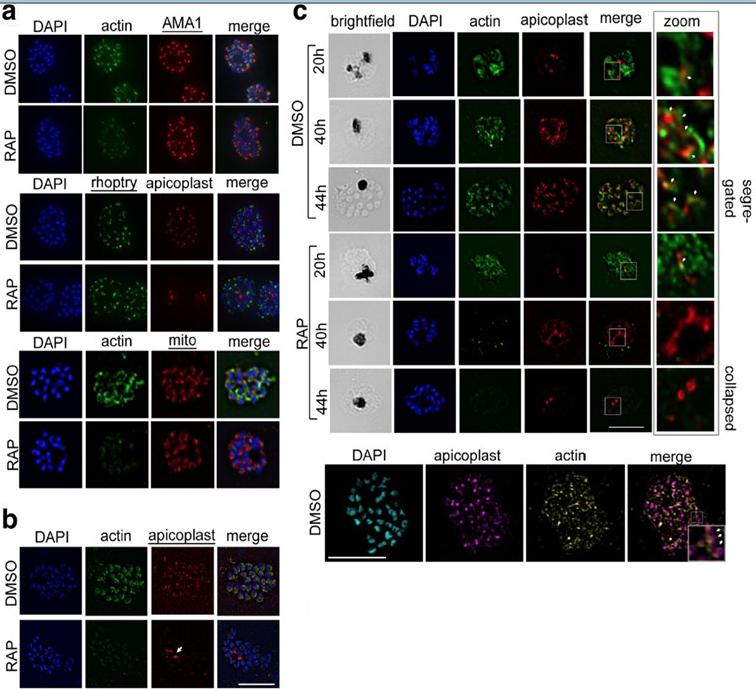
PfACT1 is not required for secretory organelle formation but is crucial for apicoplast segregation. a RAP-treated PfACT1 KO parasites have similar microneme (anti-AMA1), rhoptry (anti-RhopH2) and mitochondria (mito; anti-TOM40) architecture as DMSO controls, as revealed by IFA. b The apicoplast in PfACT1 KO parasites fails to segregate to daughter merozoites and collapses to an amorphous mass close to the food vacuole (white arrow). c IFA of samples drawn at various time points shows that apicoplast reticulation and division increase with nuclear division (DMSO, 20, 40, 44 h). The apicoplast shows close apposition to F-actin staining (zoom, white arrows). Actin staining disappears within 40 h of RAP treatment. The apicoplast does not show reticulation and extensive migration in the absence of PfACT1 (RAP, 40 and 44 h). Scale bar 5 μm. Bottom panel: Super-resolution imaging reveals close apposition of apicoplasts on the actin network. Enlarged inset in ‘merge’ shows apicoplast colocalised to actin filament (white arrows). Colocalisation analysis of apicoplast on actin in the entire image yielded a Manders coefficient of 0.83.Das S, Lemgruber L, Tay CL, Baum J, Meissner M. Multiple essential functions of Plasmodium falciparum actin-1 during malaria blood-stage development. BMC Biol. 2017 Aug 15;15(1):70.
See original on MMPMore information
| PlasmoDB | PVX_099930 |
| GeneDB | PVX_099930 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | Pv099930 |
| Orthologs | PBANKA_0830200 , PCHAS_0830500 , PF3D7_0929400 , PKNH_0727900 , PVP01_0727900 , PY17X_0833500 |
| Google Scholar | Search for all mentions of this gene |