PVX_083260 nucleoside transporter 1, putative (NT1)
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. berghei ANKA |
Possible |
RMgm-836 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-831 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-387 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-5292 | Imported from RMgmDB | |
| P. falciparum 3D7 |
Possible |
16751273 Only viable under nutrient-supplemented conditions, auxotrophic for hypoxanthine, inosine, and adenosine |
Theo Sanderson, Wellcome Trust Sanger Institute | |
| P. falciparum 3D7 |
Refractory |
USF piggyBac screen (Insert. mut.) | USF PiggyBac Screen | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-836
Reduced growth and multiplication of asexual blood stage parasites. Parasites do not induce experimental cerebral complications in C57BL/6 mice and mice can resolve infections. |
Imported from RMgmDB |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-831
Blood stages show reduced growth/multiplication rate. Parasites do not induce experimental cerebral malaria (ECM) in C57Bl/6 mice. These mice die from high parasitemias. Balb/C mice develop infections with hyperparasitemias (>50%). |
Imported from RMgmDB |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-387
Asexual blood stages show a strongly reduced growth and multiplication. BALB/c mice infected with 50 Pynt1- did not show detectable parasitemia by thin blood smears up to 30 days p.i. Mice infected with 5000 Pynt1- became patent at day 8 p.i. with a peak average parasitemia of 2.4 % at day 12 p.i.In contrast, mice infected with 50 and 5000 wild-type BS parasites became patent at days 4 and 2 p.i., respectively, and showed average peak parasitemias of 30.2% and 36.5% at days 12 and 11 p.i., respectively. |
Imported from RMgmDB |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-5292
(some) evidence presented for reduced blood stage growth/multiplication and reduced/absence of virulence |
Imported from RMgmDB |
| P. falciparum 3D7 | Asexual |
Attenuated |
16751273 Only viable under nutrient-supplemented conditions, auxotrophic for hypoxanthine, inosine, and adenosine |
Theo Sanderson, Wellcome Trust Sanger Institute |
| P. berghei ANKA | Gametocyte |
Difference from wild-type |
RMgm-387
It is reported that mutant parasites show an inefficient micro- and macrogametocytogenesis and a delayed male gamete exflagellation (however, no data is shown). |
Imported from RMgmDB |
| P. berghei ANKA | Oocyst |
Difference from wild-type |
RMgm-387
No oocyst formation |
Imported from RMgmDB |
| P. berghei ANKA | Sporozoite |
Difference from wild-type |
RMgm-387
No oocyst and sporozoite formation |
Imported from RMgmDB |
| P. berghei ANKA | Liver |
No difference |
RMgm-387 | Imported from RMgmDB |
Imaging data (from Malaria Metabolic Pathways)
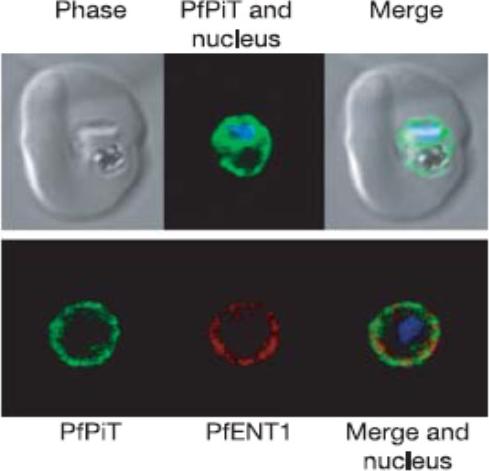
A P. falciparum-infected erythrocyte expressing PfPiT-HA (green) and nucleus stained with Hoechst 33258 (blue). Within the infected cell, PfPiT-HA localizes to the parasite. Lower row of three panels: PfPiT localizes to the parasite surface, co-localizing with PfENT1 (red), a transport protein shown previously to reside on the plasma membrane of the parasite.Saliba KJ, Martin RE, Bröer A, Henry RI, McCarthy CS, Downie MJ, Allen RJ, Mullin KA, McFadden GI, Bröer S, Kirk K. Sodium-dependent uptake of inorganic phosphate by the intracellular malaria parasite. Nature. 2006 443:582-5.
See original on MMP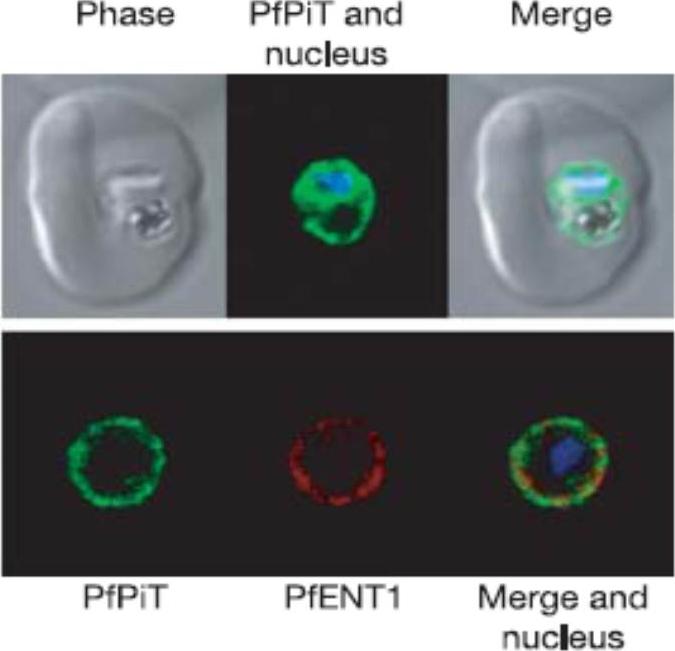
A P. falciparum-infected erythrocyte expressing PfPiT-HA (green) and nucleus stained with Hoechst 33258 (blue). Within the infected cell, PfPiT-HA localizes to the parasite. Lower row of three panels: PfPiT localizes to the parasite surface, co-localizing with PfENT1 (red), a transport protein shown previously to reside on the plasma membrane of the parasite.Saliba KJ, Martin RE, Bröer A, Henry RI, McCarthy CS, Downie MJ, Allen RJ, Mullin KA, McFadden GI, Bröer S, Kirk K. Sodium-dependent uptake of inorganic phosphate by the intracellular malaria parasite. Nature. 2006 443:582-5
See original on MMP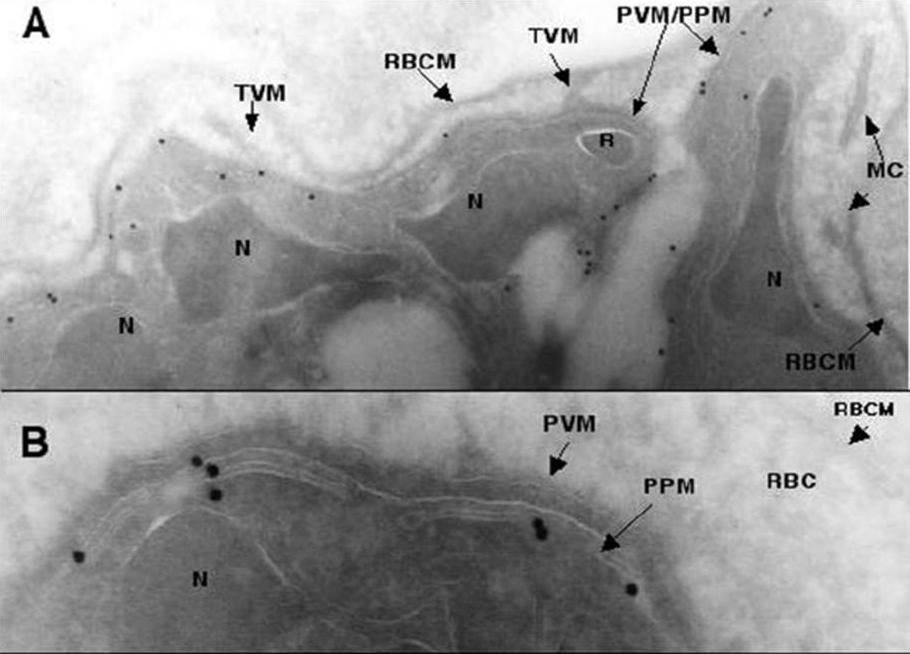
Transmission electron micrographs of ultrathin cyrosections of P. falciparum intraerythrocytic schizont stages using PfNT1 antibodies. Immunogold labeling (15 nm gold particles) of PfNT1 antibodies bound to intraerythrocytic P. falciparum is depicted. Magnification is x 12,000 in panel A and x 50,000 in panel B. MC, Maurer’s clefts; N, nucleus; R, rhoptry; TVM, tubovesicular membrane; RBCM, red blood cell membrane; PVM, parasitophorous vacuole; PPM, parasite plasma membrane.Rager N, Mamoun CB, Carter NS, Goldberg DE, Ullman B. Localization of the Plasmodium falciparum PfNT1 nucleoside transporter to the parasite plasma membrane. J Biol Chem. 2001 276:41095-9.
See original on MMP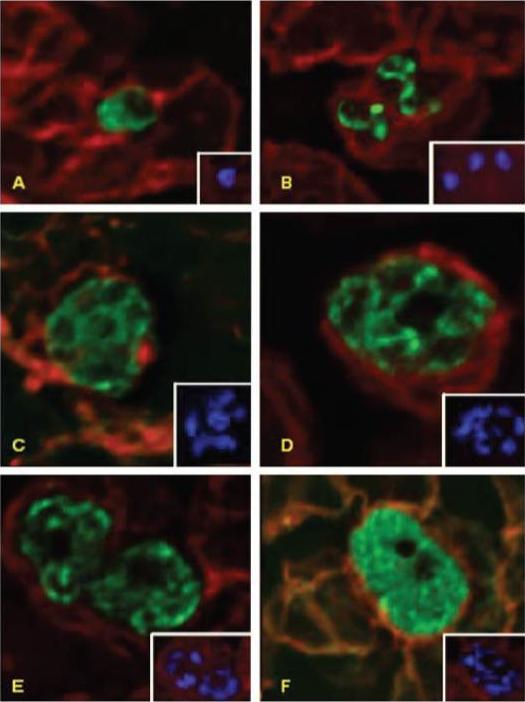
Deconvolution immunofluorescence microscopy of PfNT1 in P. falciparum-infected red blood cells. Parasites were examined by deconvolution microscopy using illumination at 546 nm to visualize PfNT1 (panels A–D), PfHT1 (panel E), and P. falciparum protein phosphatase 2C (panel F) conjugated to the FITC-conjugated anti-rabbit secondary antibody or at 488 nm to visualize Band 3 complexed with the Texas Red-conjugated anti-mouse secondary antibody (all panels). Panel A depicts a ring stage parasite, panel B shows two trophozoites, and schizonts are exhibited in panels C–F. Each panel is accompanied by an inset displaying nuclear staining with Hoechst dye.Rager N, Mamoun CB, Carter NS, Goldberg DE, Ullman B. Localization of the Plasmodium falciparum PfNT1 nucleoside transporter to the parasite plasma membrane. J Biol Chem. 2001 276:41095-9.
See original on MMP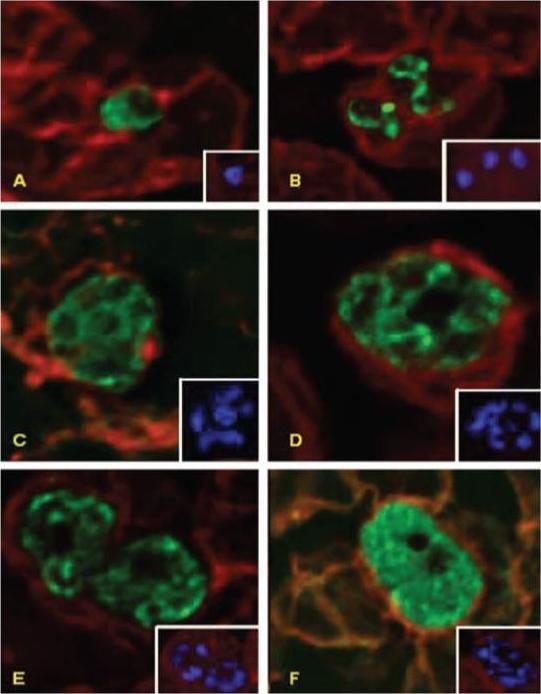
Deconvolution immunofluorescence microscopy of PfNT1 in P. falciparum-infected red blood cells. Parasites were examined by deconvolution microscopy using illumination at 546 nm to visualize PfNT1 (panels A–D), PfHT1 (panel E), and P. falciparum protein phosphatase 2C (panel F) conjugated to the FITC-conjugated anti-rabbit secondary antibody or at 488 nm to visualize Band 3 complexed with the Texas Red-conjugated anti-mouse secondary antibody (all panels). Panel A depicts a ring stage parasite, panel B shows two trophozoites, and schizonts are exhibited in panels C–F. Each panel is accompanied by an inset displaying nuclear staining with Hoechst dye.PfNT1 and PfHT1 are localized to the parasite plasma membrane. Antibodies specific for the cytosolic P. falciparum protein phosphatase 2C enzyme produce a diffuse staining pattern indicative of its cytoplasmic milieu.Rager N, Mamoun CB, Carter NS, Goldberg DE, Ullman B. Localization of the Plasmodium falciparum PfNT1 nucleoside transporter to the parasite plasma membrane. J Biol Chem. 2001 276:41095-9.
See original on MMP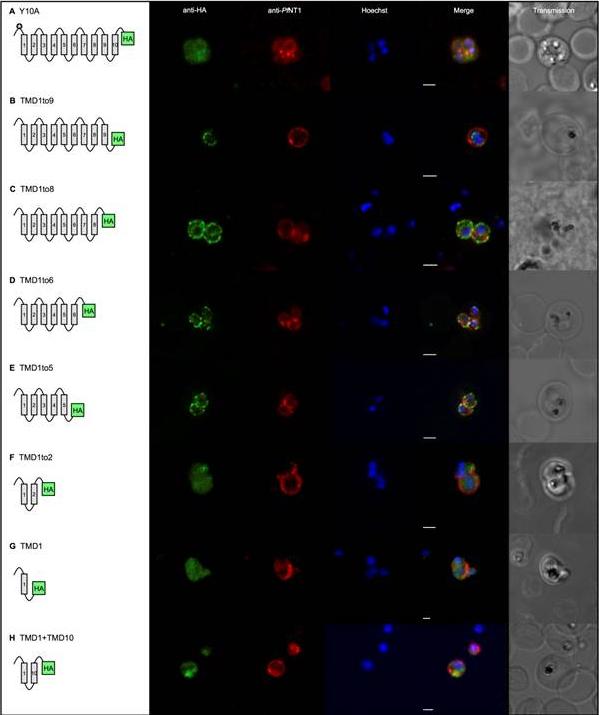
Gene expression constructs not targeted to the apicoplast now co-localised with the plasma membrane. TMDs are represented by boxes and are joined by loops. All PfoTPT constructs are episomally expressed under the PfCRT promoter and tagged with triple HA at the C-terminus detected with anti-HA and secondary antibody conjugated to FITC (green). The parasite plasma membrane is detected with anti-PfNTP1 (red). Nuclei are stained with Hoechst (blue), and transmitted light images of the parasites within their host red blood cell are shown at right. Scale bars = 2μm. A. Point mutation of tyrosine residue at position 10 (✪) in the synthetic PfoTPT (Y10A) relocates protein from the apicoplast (B) to the plasma membrane. B. Removal of TMD 10 (TMD1to9), which completely abrogates targeting to the apicoplast (C), does not result in plasma membrane localisation. C. Removal of TMDs 9 and 10 (TMD1to8), which completely abrogates targeting to the apicoplast (D), results in some protein localisation to the plasma membrane. D. Removal of TMDs 7, 8, 9 and 10 (TMD1to6), which completely abrogates targeting to the apicoplast (E), results in localisation of the protein that mostly overlaps with the parasite plasma membrane marker, PfNT1. Note that the erythrocyte is infected with three separate parasites. E. Removal of TMDs 6, 7, 8, 9 and 10 (TMD1to5), which completely abrogates targeting to the apicoplast (F), results in localisation of the protein that mostly overlaps with the parasite plasma membrane marker, PfNT1. Note that the erythrocyte is infected with two separate parasites. F. Removal of TMDs 3, 4, 5, 6, 7, 8, 9 and 10 (TMD1to2), which completely abrogates targeting to the apicoplast (G), results in some re-localisation of the protein to the plasma membrane. G. Removal of TMDs 2, 3, 4, 5, 6, 7, 8, 9 and 10 (TMD1), which completely abrogates targeting to the apicoplast (H), results in some localisation of the protein to the plasma membrane. H. Recombining TMD1 with TMD10 (TMD1+10) was unable to reconstitute apicoplast targeting (I) and resulted in some plasma membrane targeting.Lim L, Sayers CP, Goodman CD, McFadden GI. Targeting of a Transporter to the Outer Apicoplast Membrane in the Human Malaria Parasite Plasmodium falciparum. PLoS One. 2016 11(7):e0159603.
See original on MMP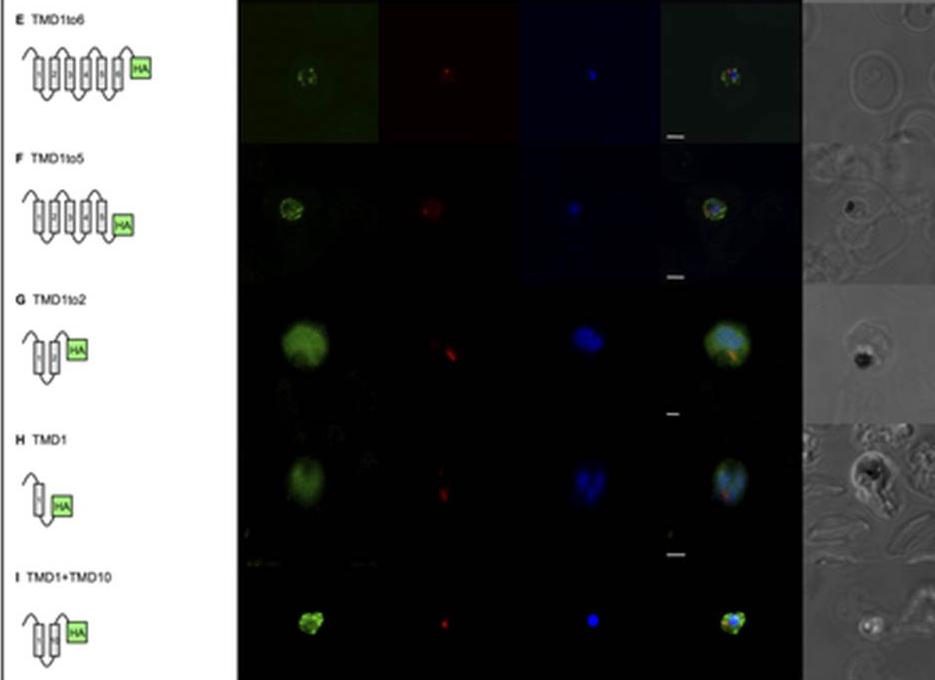
Gene expression constructs not targeted to the apicoplast now co-localised with the plasma membrane. Removal of TMDs 7, 8, 9 and 10 (TMD1to6), which completely abrogates targeting to the apicoplast (Fig 2E), results in localisation of the protein that mostly overlaps with the parasite plasma membrane marker, PfNT1. Note that the erythrocyte is infected with three separate parasites. E. Removal of TMDs 6, 7, 8, 9 and 10 (TMD1to5), which completely abrogates targeting to the apicoplast (Fig 2F), results in localisation of the protein that mostly overlaps with the parasite plasma membrane marker, PfNT1. Note that the erythrocyte is infected with two separate parasites. F. Removal of TMDs 3, 4, 5, 6, 7, 8, 9 and 10 (TMD1to2), which completely abrogates targeting to the apicoplast (Fig 2G), results in some re-localisation of the protein to the plasma membrane. G. Removal of TMDs 2, 3, 4, 5, 6, 7, 8, 9 and 10 (TMD1), which completely abrogates targeting to the apicoplast (Fig 2H), results in some localisation of the protein to the plasma membrane. H. Recombining TMD1 with TMD10 (TMD1+10) was unable to reconstitute apicoplast targeting (Fig 2I) and resulted in some plasma membrane targeting.Lim L, Sayers CP, Goodman CD, McFadden GI. Targeting of a Transporter to the Outer Apicoplast Membrane in the Human Malaria Parasite Plasmodium falciparum. PLoS One. 2016 11(7):e0159603.
See original on MMP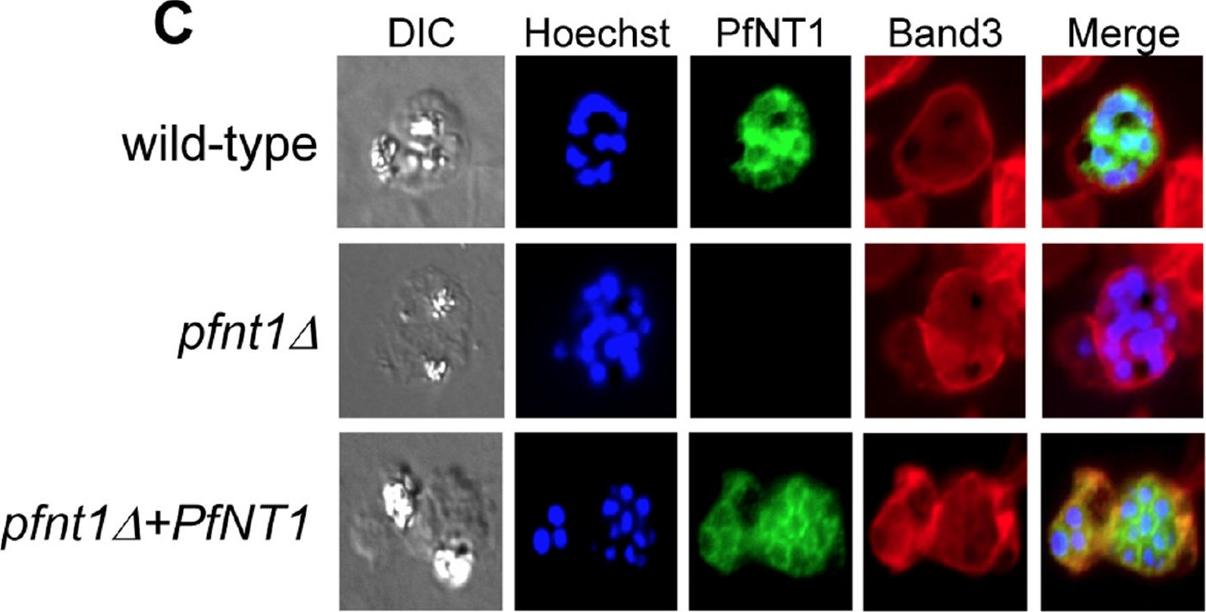
Immunofluorescence microscopy of red blood cells infected with wild-type, pfnt1Δ or pfnt1Δ+PfNT1 (PfNT1 gene was expressed episomally within the pfnt1Δ genetic background) schizont stage parasites. PfNT1 is visualized in green, Band-3 in red and nuclei in blue.El Bissati K, Downie MJ, Kim SK, Horowitz M, Carter N, Ullman B, Ben Mamoun C. Genetic evidence for the essential role of PfNT1 in the transport and utilization of xanthine, guanine, guanosine and adenine by Plasmodium falciparum. Mol Biochem Parasitol. 2008 Oct;161(2):130-9. P
See original on MMPMore information
| PlasmoDB | PVX_083260 |
| GeneDB | PVX_083260 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | Pv083260 |
| Orthologs | PBANKA_1360100 , PCHAS_1364700 , PF3D7_1347200 , PKNH_1253700 , PVP01_1207600 , PY17X_1365800 |
| Google Scholar | Search for all mentions of this gene |