PVP01_1212200 myosin A, putative
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. berghei ANKA |
Refractory |
PlasmoGEM (Barseq) | PlasmoGEM | |
| P. falciparum 3D7 |
Refractory |
USF piggyBac screen (Insert. mut.) | USF PiggyBac Screen | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. falciparum 3D7 | Asexual |
Attenuated |
31337750 Almost complete lack of merozoites after invasion assay, which is interpreted as an invasion defect. (Although an egress defect not ruled out by this work as far as I can see) |
Theo Sanderson, Francis Crick Institute |
Imaging data (from Malaria Metabolic Pathways)
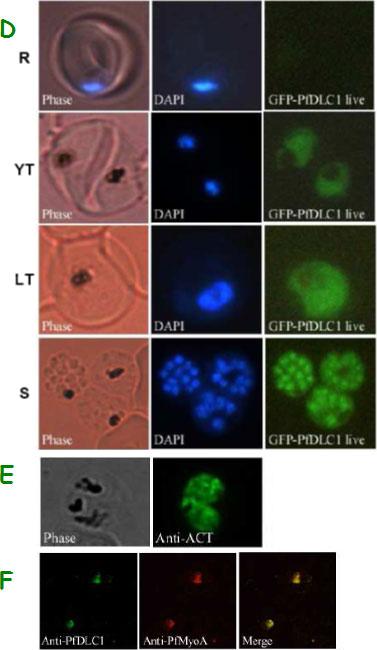
Expression of PfDLC1, PfACT1, and PfMyoA gene products by wild type and transfected P. falciparum. D. Expression and localization of PfDLC1-GFP throughout the erythrocytic cell cycle of P. falciparum. Live transfected parasites were analysed by fluorescence microscopy. E. Immunofluorescence localization of P. falciparum actin in fixed transgenic parasites (late trophozoites). F. Co-localization of PfDLC1 and PfMyoA. Smears of erythrocytes infected with 3D7 strain were fixed and probed with rat anti-PfDLC1 followed by anti-rat IgG-FITC and rabbit anti-PfMyoA, followed by anti-rabbit IgG-Alexa Fluor 568 (red). PfDLC1 location showed a homogenous distribution in the cytoplasm of the parasite (D). It seems that the protein is not exported to the nucleus of P. falciparum or to the RBC cytoplasm or to the RBC surface (Fig. 7C and 7D). The expression of PfDLC1 in younger stage transfectant as well as in the wild type stage (young ring) was undetectable (D). P. falciparum actin is distributed in the parasite cytoplasm (E). Myosin A partially overlap PfDLC1 in schizonts (F).Daher W, Pierrot C, Kalamou H, Pinder JC, Margos G, Dive D, Franke-Fayard B, Janse CJ, Khalife J. Plasmodium falciparum dynein light chain 1 interacts with actin/myosin during blood stage development. J Biol Chem. 2010 285:20180-20191.
See original on MMP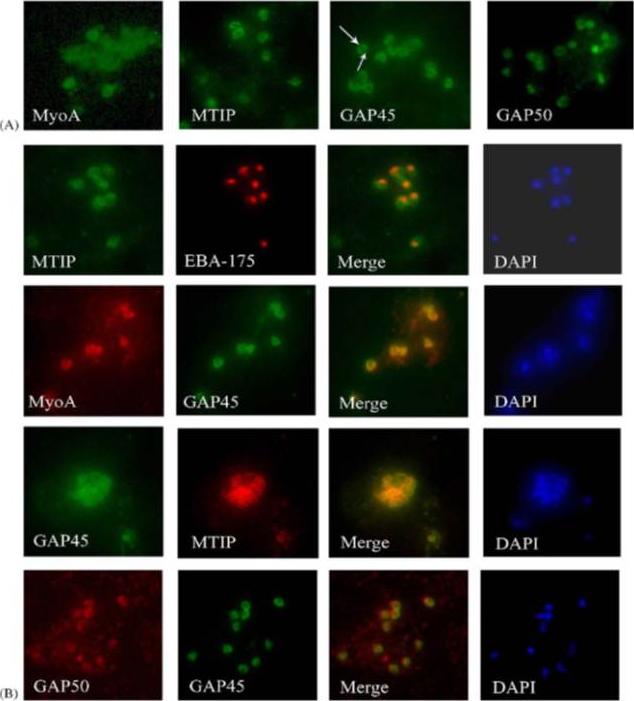
Components of the P. falciparum merozoite glideosome localize to the periphery of the parasite. (A) Indirect-immunofluorescence of individual glideosome components in late blood-stage parasites: Thin smears of P. falciparum infected erythrocytes were probed with anti-glideosome sera. The arrow indicates a gap in peripheral staining at the end of the merozoite, suggestive of an inner membrane complex (IMC) localization. (B) Indirect-immunofluorescence of late blood-stage parasites co-stained with antisera to other glideosome components and PfEBA-175: Glideosome antisera were used in co-immunofluorescence assays to assess co-localization. Anti-PfEBA-175 antisera recognize an invasion ligand that is localized to the micronemes, and is used as a marker for the apical end of the merozoite.Jones ML, Kitson EL, Rayner JC. Plasmodium falciparum erythrocyte invasion: a conserved myosin associated complex. Mol Biochem Parasitol. 2006 147:74-84. Copyright Elsevier 2009.
See original on MMP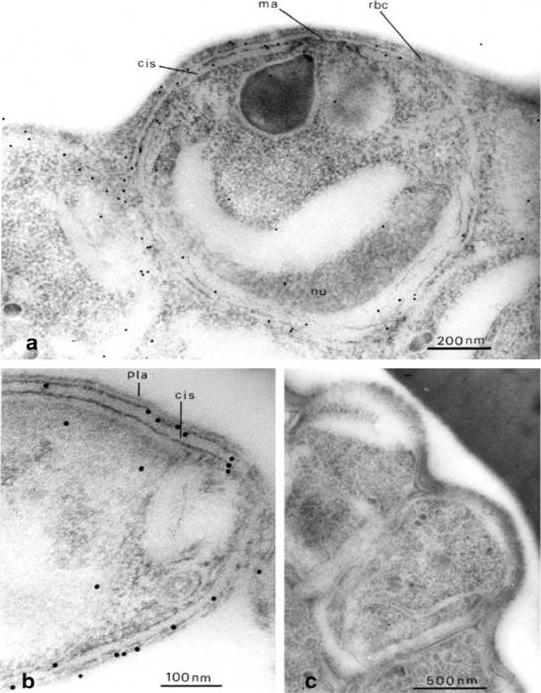
Immuno-electron microscopy, showing distribution of Pf-myo1 in the malaria parasite. (a) Immunogold-labelled merozoites before their release from the red cell. The parasite was labelled with anti-Pf-myo1 and second antibody, conjugated with 10 nm gold particles. The myosin is located around the periphery of the merozoite. In the centre the merozoite plasma membrane overlies the apical prominence (ma) close to the large pearshaped secretory rhoptry, and elsewhere in the parasite a flat cisterna (cis) underlies the plasma membrane. The curved nucleus (nu) is visible at the base of the merozoite. Note absence of label from the flat tip of the apical prominence, and the greater frequency of labelling around the apical prominence than elsewhere. (b) Longitudinal/oblique section through the apical end of a free merozoite. Labelling is associated with the cortical cytoplasm between the plasma membrane (pla) and the outer membrane of the subplasmalemmal cisterna (cis). (c) Control section through a group of mature merozoites at the periphery of a late-stage schizont, incubated with preimmune serum before gold labelling. No significant labelling can be observed.Adapted with permission from Pinder JC, Fowler RE, Dluzewski AR, Bannister LH, Lavin FM, Mitchell GH, Wilson RJ, Gratzer WB. Actomyosin motor in the merozoite of the malaria parasite, Plasmodium falciparum: implications for red cell invasion. J Cell Sci. 1998 111:1831-9.
See original on MMP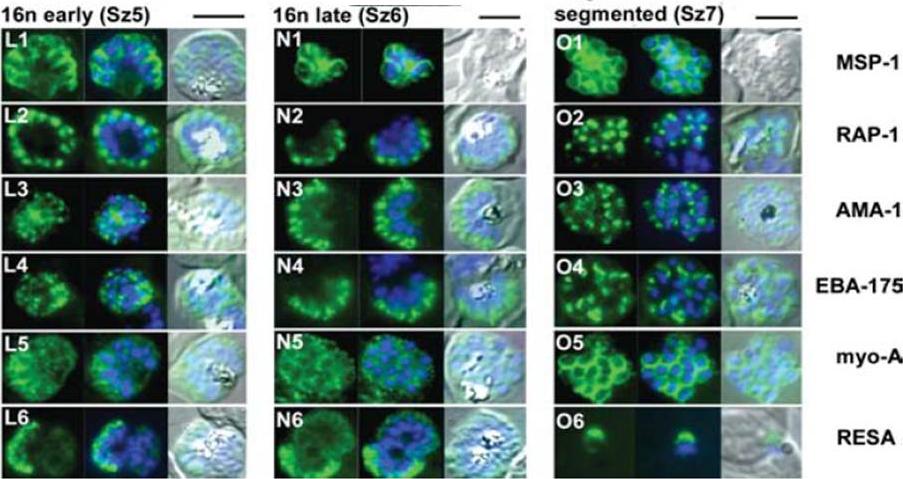
The appearance of Plasmodium falciparum schizonts of the ITO4 line at different times of development. The time of sampling from invasion and the number of nuclei per schizont are indicated. Antibodies to proteins associated with invasion related organelles or the parasite periphery (rhoptries : RAP-1; micronemes: AMA-1, EBA-175; dense granules: RESA; parasite periphery: MSP-1, myo-A) were chosen to follow the development and assembly of merozoites in maturing schizonts.Margos G, Bannister LH, Dluzewski AR, Hopkins J, Williams IT, Mitchell GH. Correlation of structural development and differential expression of invasion-related molecules in schizonts of Plasmodium falciparum. Parasitology. 2004 129:273-87. Copyright Cambridge University Press Journals 2011
See original on MMP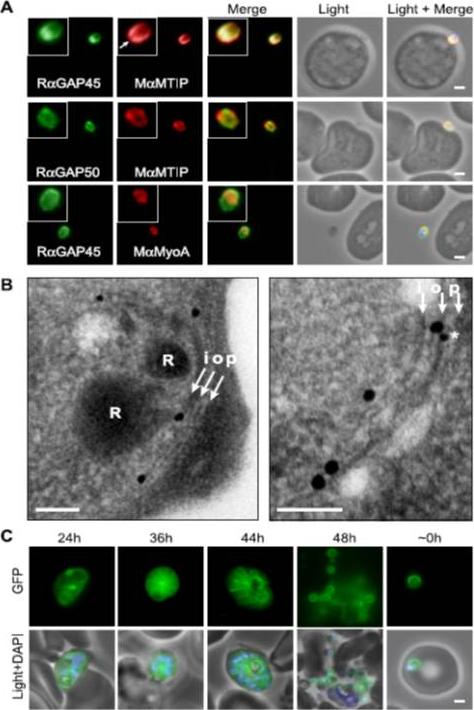
Localization of PfGAP proteins to the inner membrane complex. A, PfGAP50 and PfGAP45 co-localize with MTIP (a protein known to localize to the inner membrane complex) and partially with PfMyoA (known to associate with the innermembrane complex and merozoite apical tip (6, 38)). The white arrow in the MTIP inset points to an area of reduced fluorescence at the apical tip in a single z-stack, consistent with the absence of IMC at this position. B, immunoelectron microscopy confirms IMC localization for PfGAP45 (large 25-nm gold particle). The white arrows indicate trilaminar appearance of the IMC and merozoite plasma membrane. i, inner IMC membrane; o, outer IMC membrane; p, plasma membrane R, rhoptry. *, single anti-PfGAP50-conjugated gold particle found in close proximity to PfGAP45 but giving poor reactivity (typically 1 particle/merozoite). Scale bar in electron microscopy, 0.2 mm. C, PfGAP45-GFP shows fluorescence that concentrates to the merozoite periphery late in asexual development, consistent with the Scale bar in IFA, 1 mm.Baum J, Richard D, Healer J, Rug M, Krnajski Z, Gilberger TW, Green JL, Holder AA, Cowman AF. A conserved molecular motor drives cell invasion and gliding motility across malaria life cycle stages and other apicomplexan parasites. J Biol Chem. 2006 281(8):5197-208.
See original on MMP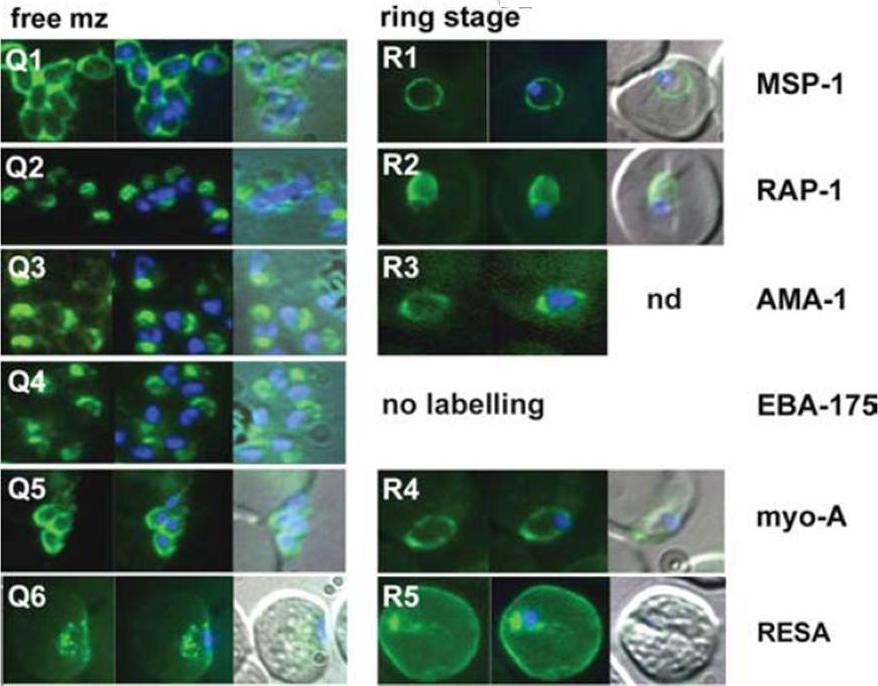
The appearance of Plasmodium falciparum merozoites and rings of the ITO4 line at different times of development. Antibodies to proteins associated with invasion related organelles or the parasite periphery (rhoptries : RAP-1; micronemes: AMA-1, EBA-175; dense granules: RESA; parasite periphery: MSP-1, myo-A) were chosen to follow the development and assembly of merozoites and rings.Margos G, Bannister LH, Dluzewski AR, Hopkins J, Williams IT, Mitchell GH. Correlation of structural development and differential expression of invasion-related molecules in schizonts of Plasmodium falciparum. Parasitology. 2004 129:273-87. Copyright Cambridge University Press Journals 2011
See original on MMP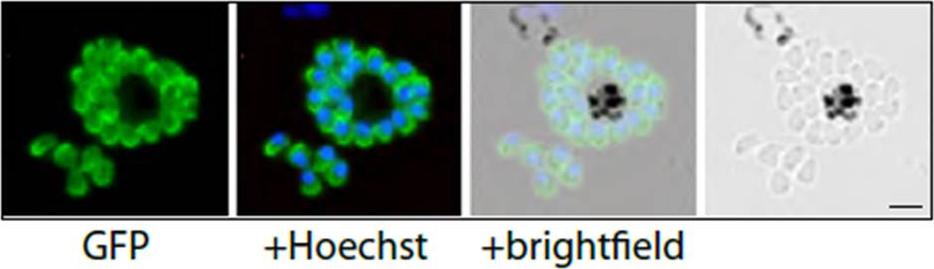
PfMyoA-GFP-expressing merozoites as viewed by live fluorescence microscopy. GFP was detected by green fluorescence and the nuclei (blue) were labelled with Hoechst dye prior to microscopic analysis; the GFP signal is distributed to the parasite periphery. Scale bar is 2 μm.Yusuf NA, Green JL, Wall RJ, Knuepfer E, Moon RW, Schulte-Huxel C, Stanway RR, Martin SR, Howell SA, Douse CH, Cota E, Tate EW, Tewari R, Holder AA. The Plasmodium Class XIV Myosin, MyoB has a Distinct Subcellular Location in Invasive and Motile Stages of the Malaria Parasite, and an Unusual Light Chain. J Biol Chem. 2015 Mar 23.. [Epub ahead of print]
See original on MMP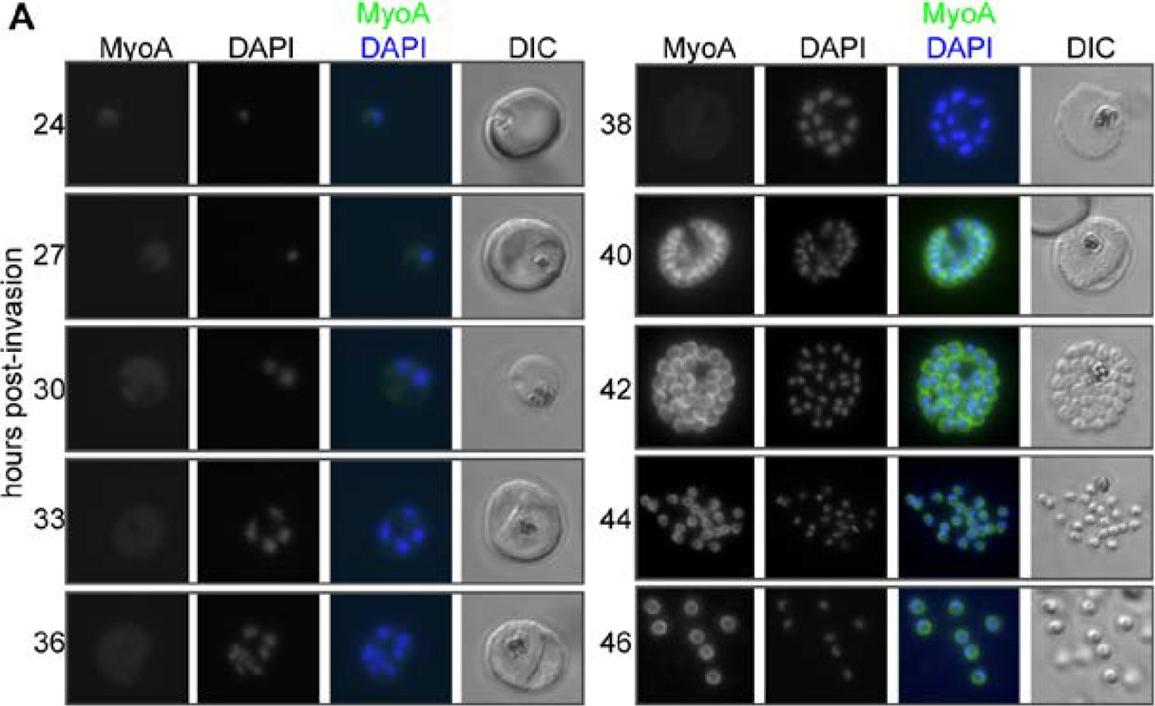
MyoA-GFP expression and glideosome complex formation during the late stages of intracellular development of P. falciparum in the red blood cell. (A) MyoA-GFP expression was detected by live fluorescence microscopy and the nuclei were detected by staining with DAPI. The merged colour image with MyoA-GFP (green) and DAPI (blue) and differential interference contrast (DIC) bright field images are also shown. Schizogony starts at around 30 hours post-invasion and MyoA-GFP is detected from 38–40 hours in multinucleated forms. Scale bar: 2 µm. MyoA-GFP was largely undetectable by live fluorescence microscopy until approximately 38 hours after merozoite invasion of an erythrocyte, but was present in later stages at the periphery of developing segmented schizonts and free merozoites (but not associated with the food vacuole/residual body), consistent with its proposed location at the IMC (Fig. 2A). No GFP signal was detected in ring or trophozoite stages indicating that the protein is not synthesised at this time and any MyoA carried through with the invading merozoite is degraded rapidly following erythrocyte invasion.Green JL, Wall RJ, Vahokoski J, Yusuf NA, Ridzuan MAM, Stanway RR, Stock J, Knuepfer E, Brady D, Martin SR, Howell SA, Pires IP, Moon RW, Molloy JE, Kursula I, Tewari R, Holder AA. Compositional and expression analyses of the glideosome during the Plasmodium life cycle reveal an additional myosin light chain required for maximum motility. J Biol Chem. 2017 Sep 11.
See original on MMP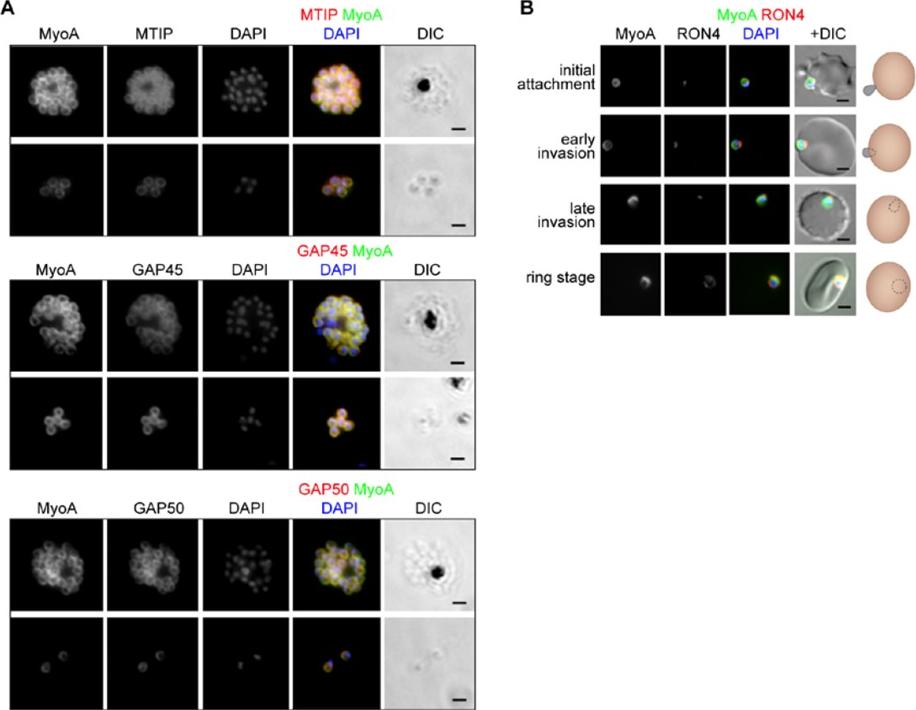
The location of MyoA during P. falciparum merozoite invasion. (A) MyoA-GFP is located at the periphery of developing intracellular (top row of each pair of images) and free extracellular merozoites (bottom row), and is colocalized with antibodies specific for IMC proteins MTIP, GAP45 and GAP50. In the merged colour image the MyoA-GFP signal is green and antibodies specific for the IMC proteins are red; nuclei are stained blue with DAPI. The DIC image is also shown. (B) Individual merozoites are captured at different stages of invasion from initial attachment, through early and late invasion to the intracellular ring stage. MyoA-GFP remains peripheral whereas RON4, initially in the apical rhoptry neck, relocates during invasion. Merged colour images with MyoA-GFP (green), RON4 (red), and nuclei (blue) and DIC images are also shown, together with a schematic of each cell-pair. Scale bar: 2 μm.Green JL, Wall RJ, Vahokoski J, Yusuf NA, Ridzuan MAM, Stanway RR, Stock J, Knuepfer E, Brady D, Martin SR, Howell SA, Pires IP, Moon RW, Molloy JE, Kursula I, Tewari R, Holder AA. Compositional and expression analyses of the glideosome during the Plasmodium life cycle reveal an additional myosin light chain required for maximum motility. J Biol Chem. 2017 Sep 11.
See original on MMPMore information
| PlasmoDB | PVP01_1212200 |
| GeneDB | PVP01_1212200 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | null |
| Orthologs | PBANKA_1355700 , PCHAS_1360300 , PF3D7_1342600 , PKNH_1258800 , PVX_083030 , PY17X_1361400 |
| Google Scholar | Search for all mentions of this gene |