PVP01_0109300 chloroquine resistance transporter, putative (CRT)
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. berghei ANKA |
Refractory |
RMgm-612 | Imported from RMgmDB | |
| P. berghei ANKA |
Refractory |
PlasmoGEM (Barseq) | PlasmoGEM | |
| P. falciparum 3D7 |
Refractory |
12813054 | Theo Sanderson, Wellcome Trust Sanger Institute | |
| P. falciparum 3D7 |
Refractory |
USF piggyBac screen (Insert. mut.) | USF PiggyBac Screen | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. falciparum 3D7 | Asexual |
Difference from wild-type |
35867395 (Knock down)
\"Here, we report on the generation of a conditional PfCRT knock-down mutant in P. falciparum. The mutant accumulated oligopeptides 2 to at least 8 residues in length under knock-down conditions, as shown by comparative global metabolomics. The accumulated oligopeptides were structurally diverse, had an isoelectric point between 4.0 and 5.4 and were electrically neutral or carried a single charge at the digestive vacuolar pH of 5.2. Fluorescently labeled dipeptides and live cell imaging identified the digestive vacuole as the compartment where oligopeptides accumulated. Our findings suggest a function of PfCRT in oligopeptide transport across the digestive vacuolar membrane in P. falciparum and associated with it a role in nutrient acquisition and the maintenance of the colloid osmotic balance.\" |
Theo Sanderson, Francis Crick Institute |
Imaging data (from Malaria Metabolic Pathways)
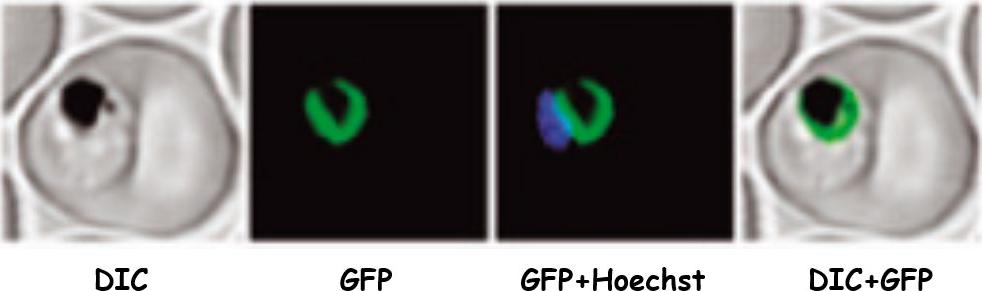
Full-length PfCRT fused to the green fluorescent protein (GFP). Confocal laser scanning microscopy of parasites expressing PfCRTfull revealed a fluorescence signal surrounding the malaria pigment, consistent with a localization of the corresponding protein at the digestive vacuolar membrane. First column, differential interference contrast (DIC); second column, GFP; third column, GFP and nuclear staining with Hoechst; fourth column overlay of green and DIC channel. Scale bar = 4 mm.Kuhn Y, Sanchez CP, Ayoub D, Saridaki T, van Dorsselaer A, Lanzer M. Trafficking of the phosphoprotein PfCRT to the digestive vacuolar membrane in Plasmodium falciparum. Traffic. 2010 11:236-49. Copyright John Wiley & Sons Ltd. 2010.
See original on MMP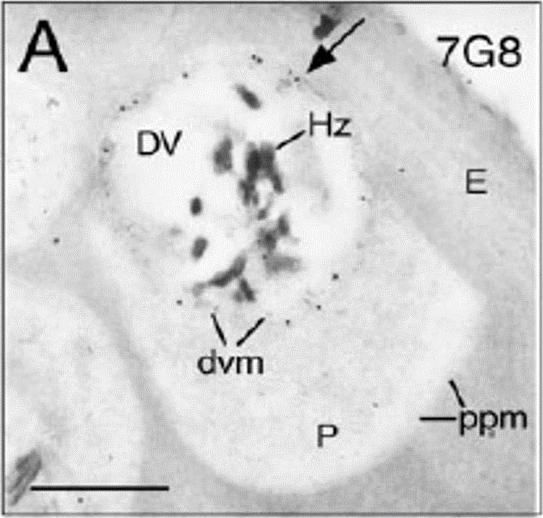
Immunoelectron microscopy was performed using affinity-purified anti-PfCRT antibodies. Representative micrographs are shown for each parasite line. Bars correspond to 0.25 mm. Arrowheads indicate immunogold-labeled PfCRT deposits on the digestive vacuole membrane (dvm). DV, digestive vacuole; E, erythrocyte; Hz, hemozoin; P, parasite; ppm, parasite plasma membrane. Waller KL, Muhle RA, Ursos LM, Horrocks P, Verdier-Pinard D, Sidhu AB, Fujioka H, Roepe PD, Fidock DA. Chloroquine resistance modulated in vitro by expression levels of the Plasmodium falciparum chloroquine resistance transporter. J Biol Chem. 2003 278:33593-601.
See original on MMP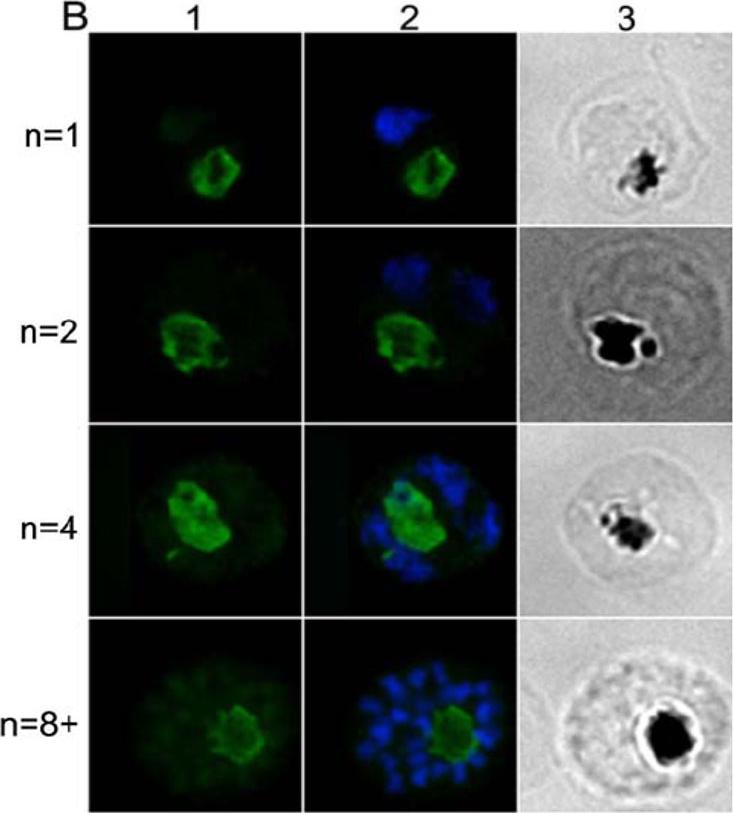
IFA location of CRT in mature parasites at different stages of nuclear division. Each row in panel B shows an identical field from 1% formaldehyde-fixed thin smears of late-stages parasites at the 1-, 2-, 4-, and 8-nuclei stage. Panel B1 shows rabbit anti-CRT antibody labeling (the secondary antibody was Alexa Fluor 488-conjugated anti-rabbit IgG antibody (green). Panel B2 shows the same fields merged with the corresponding DAPI stained nucleus images (blue). Panel B3 shows the same fields visualized by light microscopy - note the pigment is clearly visible. Rabbit-anti-CRT antiobody is clearly detectable throughout parasite maturation and is associated with the food vacuole.Dluzewski AR, Ling IT, Hopkins JM, Grainger M, Margos G, Mitchell GH, Holder AA, Bannister LH. Formation of the food vacuole in Plasmodium falciparum: a potential role for the 19 kDa fragment of merozoite surface protein 1 (MSP1(19)). PLoS ONE. 2008 Aug 3(8):e3085. ™
See original on MMP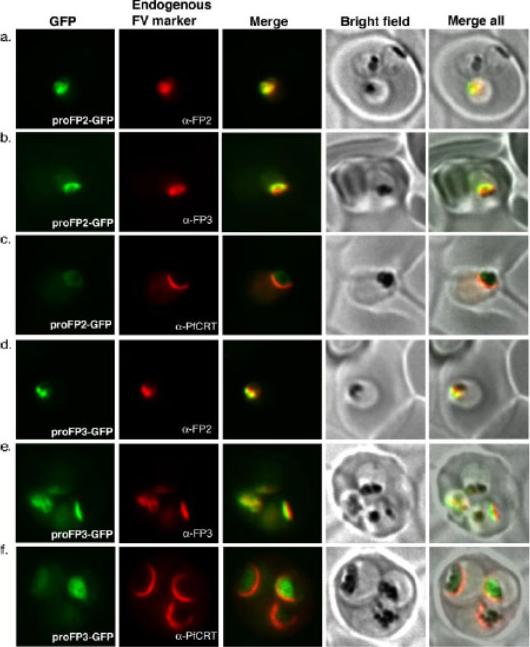
Co-localization of full-length prodomain constructs with endogenous food vacuole proteins. Trophozoites transfected with proFP2-GFP (under falcipain-2 promoter) (a–c) or proFP3-GFP (under HSP86 promoter) (d–f) constructs were processed for immunofluorescence microscopy and labeled with anti-FP2 (a,d), anti-FP3 (b, e), or anti-PfCRT.Subramanian S, Sijwali PS, Rosenthal PJ. Falcipain cysteine proteases require bipartite motifs for trafficking to the Plasmodium falciparum food vacuole. J Biol Chem. 2007 282(34):24961-9.
See original on MMP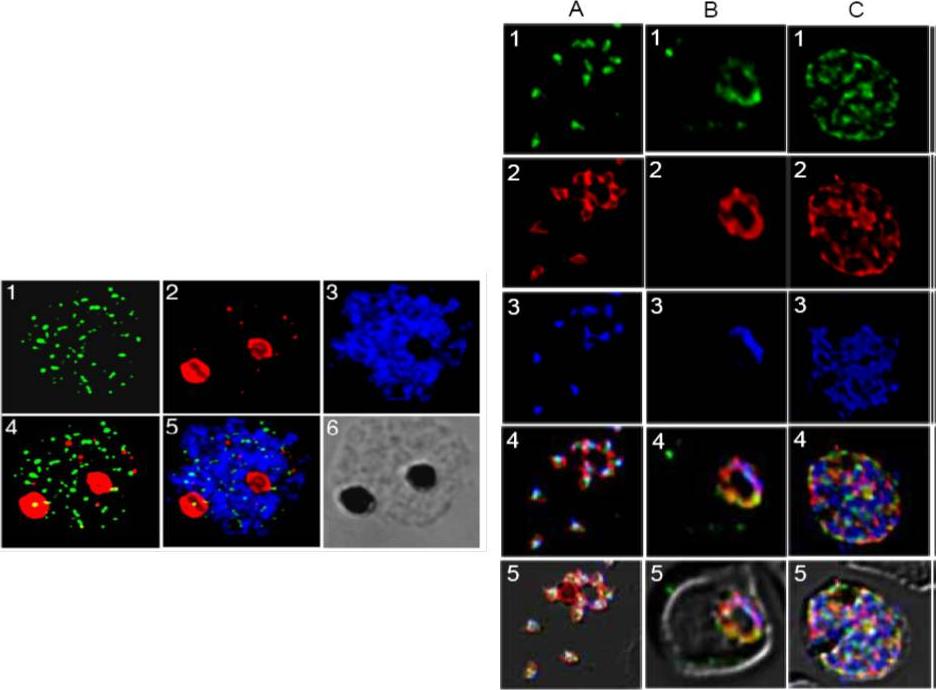
Right panel: Expression and localization of PfMRP within asexual stages of the W2 parasite. 1 (green, anti-PfMRP; 2 (red), anti-PfMSP-1 (PFI1475w); 3 (blue), DAPI dye; 4, merged 1-3; 5, merged 1-4 and diffrential interference contrast (DIC). A, free merozoites; B, a trophozoite; C, a schizont. In merozoites, PfMRP was expressed in membrane-bound vesicles located inside the parasite plasma membrane and did not co-localize with PfMSP-1 on plasma membrane. In trophozoite and schizont stages, PfMRP expression increased with parasite growth, located on an extensive network of membrane structures including plasma membrane although it appeared to be present in ‘patches’. Left panel: Immunofluorescent assay (IFA) localization of PfMRP and PfCRT in mature schizonts and merozoites. 1, anti-PfMRP; 2, anti-PfCRT, staining food vacuoles; 3, DAPI; 4, merged images of 1 and 2; 5, merged image of 1-3; and 6, differential interference contrast (DIC).Raj DK, Mu J, Jiang H, Kabat J, Singh S, Sullivan M, Fay MP, McCutchan TF, Su XZ. Disruption of a Plasmodium falciparum multidrug resistance-associated protein (PFMRP) alters its fitness and transport of antimalarial drugs and glutathione. J Biol Chem. 2008 284(12):7687-96.
See original on MMP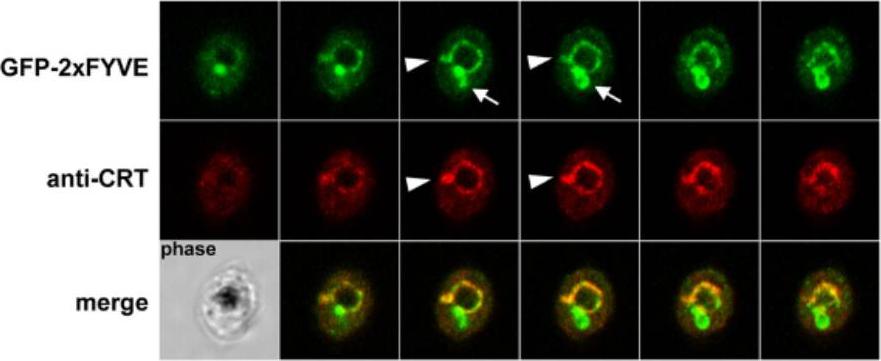
GFP fluorescence in parasites expressing GFP-2xFYVE. This construct has been used in various organisms to monitor the intracellular localization of PI3P. Colocalization of GFP-2xFYVE and the food vacuole membrane marker CRT using rabbit anti-CRT. Serial images of a Z-stack acquisition (0.38 mm step) are displayed. Arrowheads indicate protrusions from the food vacuole membrane that are colabeled with CRT and GFP-2xFYVE, while arrows indicate zones of intense GFP-2xFYVE staining in the absence of the food vacuole marker. A corresponding phase-contrast image is shown in the lower left corner. Fluorescence was associated mainly with the membrane of the food vacuole.Tawk L, Chicanne G, Dubremetz JF, Richard V, Payrastre B, Vial HJ, Roy C, Wengelnik K. Phosphatidylinositol 3-phosphate, an essential lipid in Plasmodium, localizes to the food vacuole membrane and the apicoplast. Eukaryot Cell. 2010 9:1519-30.
See original on MMP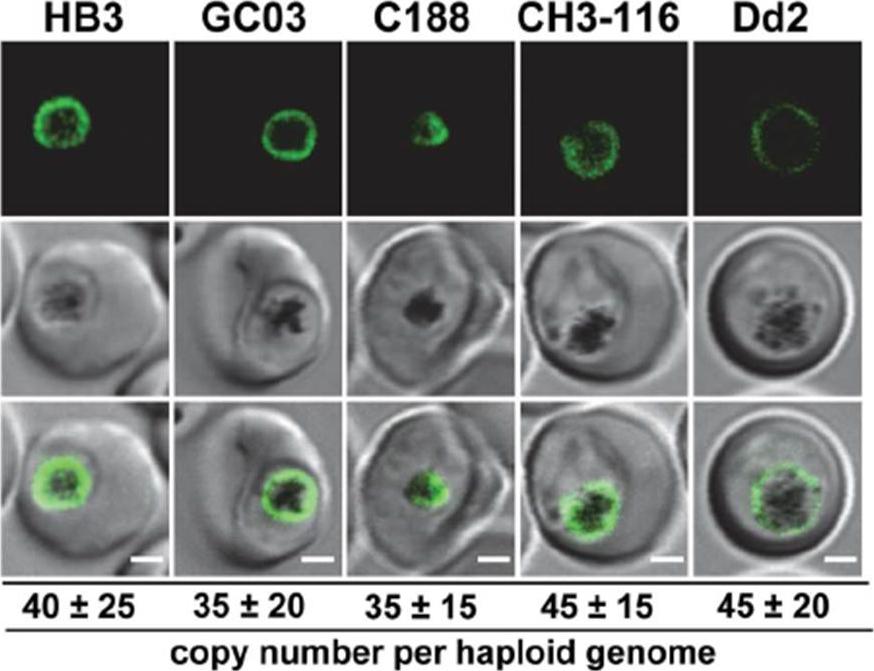
Live cell images of the P. falciparum strains indicated expressing an episomal copy of the Dd2 form of pfcrt fused to the GFP coding sequence. Fluorescence is located at the digestive vacuolar membrane, consistent with previous reports. Bar, 2 mM. The copy number of the plasmid per haploid genome is indicated. The means 6 SEM of at least 6 biological replicates are shown. The copy numbers are not statistically different in the transfectants. In all cases, the PfCRT/GFP fusion protein was expressed and localized at the membrane of the parasite’s digestive vacuole.Sanchez CP, Liu CH, Mayer S, Nurhasanah A, Cyrklaff M, Mu J, Ferdig MT, Stein WD, Lanzer M. A HECT Ubiquitin-Protein Ligase as a Novel Candidate Gene for Altered Quinine and Quinidine Responses in Plasmodium falciparum. PLoS Genet. 2014 10(5):e1004382.
See original on MMP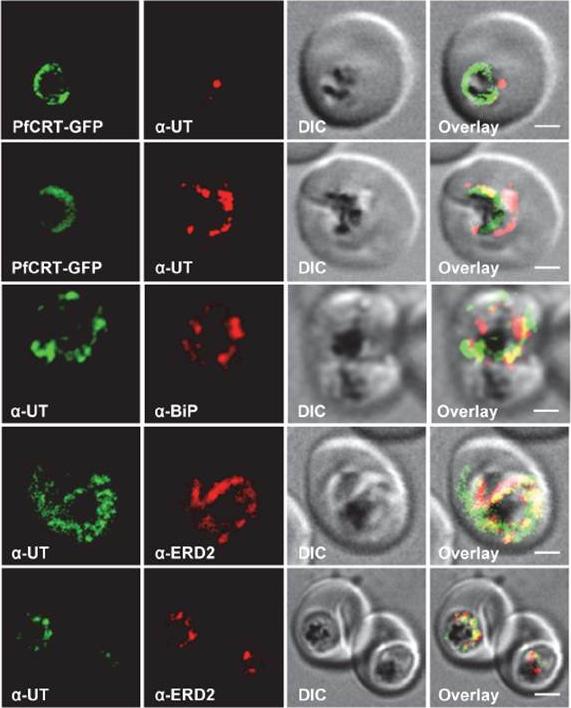
Subcellular localization of PfUT (ubiquitin-transferase). P. falciparum-infected erythrocytes at the trophozoite stage were fixed and analyzed by immunofluorescence assays using antisera to the ER marker BiP (rabbit, 1:1000), the Golgi marker ERD2 (rat, 1:500), and the N- (panels 1 and 5, rabbit, 1:3000; panel 3, mouse, 1:2000) and C-terminal domains of PfUT (panels 2 and 4, rabbit, 1:3000). Panel 1 shows a late ring stage parasite, the other panels show trophozoites. GFP fluorescence was detected, by confocal fluorescence microscopy, in parasites expressing episomally a PfCRT/GFP fusion protein. The different antisera raised against PfUT showed comparable results. Bar, 2 mm. Immunofluorescence microscopy partially co-localized PfUT with the ER marker BiP and the Golgi marker ERD2, but not with PfCRT.Sanchez CP, Liu CH, Mayer S, Nurhasanah A, Cyrklaff M, Mu J, Ferdig MT, Stein WD, Lanzer M. A HECT Ubiquitin-Protein Ligase as a Novel Candidate Gene for Altered Quinine and Quinidine Responses in Plasmodium falciparum. PLoS Genet. 2014 10(5):e1004382.
See original on MMP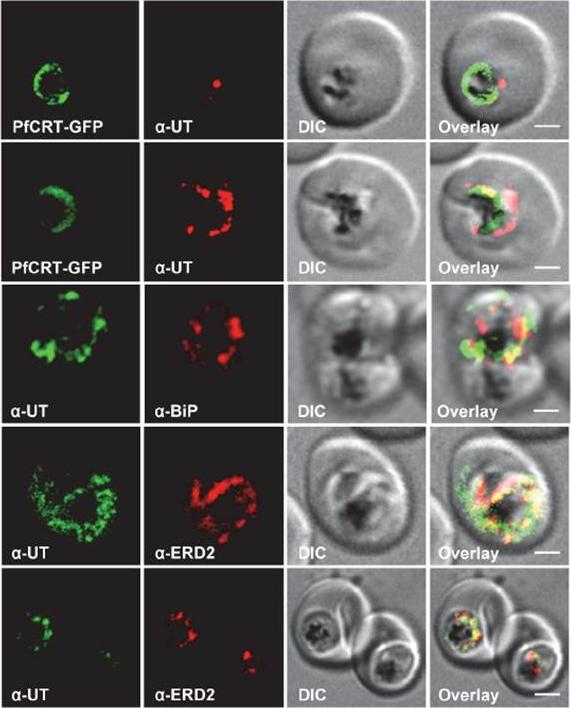
Immunofluorescence images of PfMRP2 localization labelled with Alexa488 during the development from ring to trophozoite and schizont stages in the wild-type clone 11C parasite. Nucleus was stained with DAPI (4′,6′-diamidino-2-phenylindole). The bottom panel shows dual labelling of the parasite at early schizont stage with anti-PfMRP2 (Alexa488) antibody and digestive vacuole-specific PfCRT antibody (Alexa594). In all asexual blood stages of 11C/wt, the PfMRP2 IFAsignal exhibited a punctate pattern suggesting its association with membranous vesicles within the parasite cytoplasm. In the ring stage, PfMRP2-associated compartments concentrated to the periphery of the parasite, while in trophozoite and schizont stages these were more dispersed throughout the cytoplasm. The signal also appeared discrete and punctuated around individual merozoites in the mature schizonts just prior to rupture.Mok S, Liong KY, Lim EH, Huang X, Zhu L, Preiser PR, Bozdech Z. Structural polymorphism in the promoter of pfmrp2 confers Plasmodium falciparum tolerance to quinoline drugs. Mol Microbiol. 2014 91(5):918-34.
See original on MMP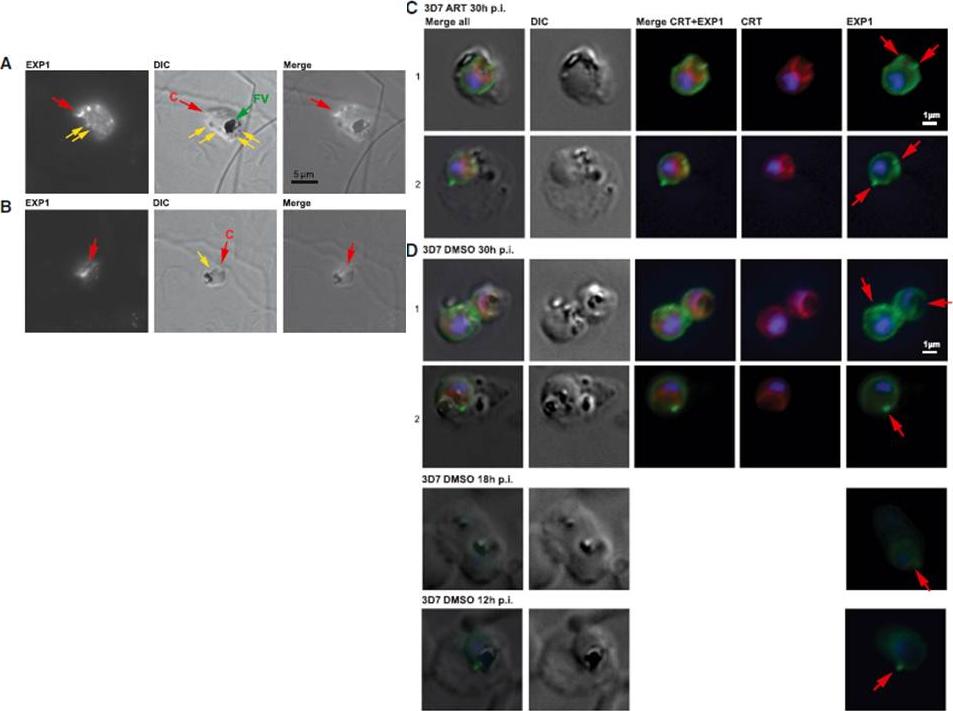
(A) Immunofluorescence microscopy of 3D7 trophozoite parasites labeled with anti-EXP1 antibodies (EXP1) shows evidence of peripheral expression of EXP1 that colocalizes with an apparent cytostomic invagination (C, red arrow; DIC panel). A fully formed trophozoite stage food vacuole (FV, green arrow) presents a large accumulation of hemozoin, whereas small darker spots (yellow arrows) might be vesicles that have pinched off from the invagination. (B) A late ring stage parasite displays a prominent spherical invagination that also partly corresponds to a convexly shaped EXP1 expression signal; small vesicles (yellow arrow) may become components of an internal pre-FV compartment (dark irregular structure). (C) Trophozoite stage 3D7 parasites exposed to ART indicate peripheral EXP1 expression (row C1) or enhanced expression foci at the PVM (row C2). (D) Control trophozoites (DMSO) displayed similar morphology in EXP1 expression patterns to the drug-exposed parasites. Antibodies to the FV marker CRT (P. falciparum chloroquine resistance marker) indicate that EXP1 expression was largely independent of FV location.Lisewski AM, Quiros JP, Ng CL, Adikesavan AK, Miura K, Putluri N, Eastman RT, Scanfeld D, Regenbogen SJ, ltenhofen L, Llinás M, Sreekumar A, Long C, Fidock DA, Lichtarge O. Supergenomic Network Compression and the Discovery of EXP1 as a Glutathione Transferase Inhibited by Artesunate. Cell. 2014 158(4):916-28.
See original on MMP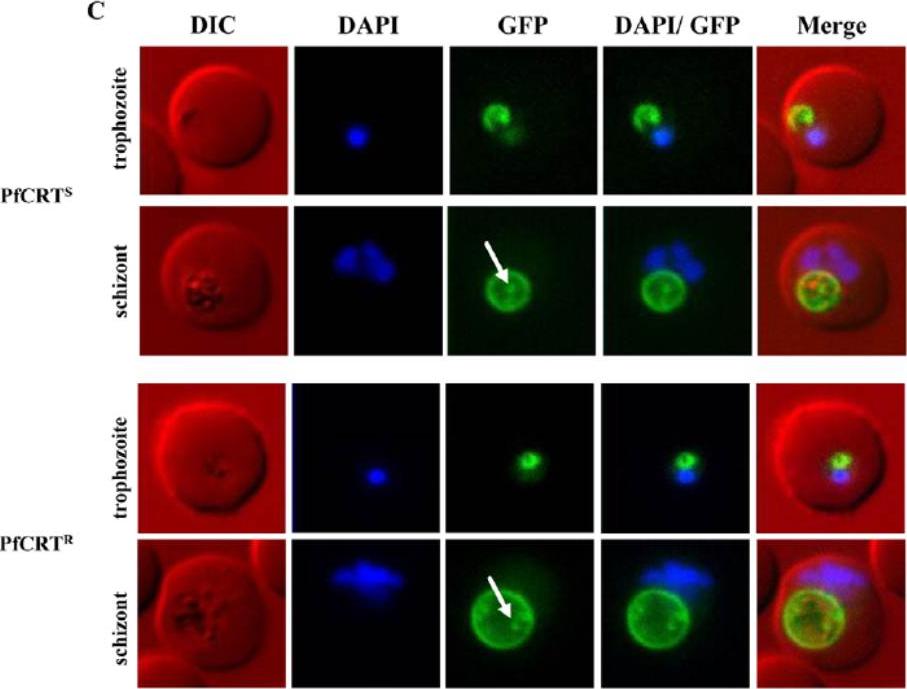
Over-expression of sensitive and resistant form of PfCRT as GFP-fusion protein. Fluorescence microscopy of GFP-fusions of sensitive (PfCRTS, top) and resistant (PfCRTR, bottom) form of PfCRT. PfCRTS and PfCRTR were over-expressed as GFP-fusion proteins using an ATc-inducible expression system. Live cell images of DAPI-stained infected red blood cells show that both forms of PfCRT localise to the FV membrane. Later stages of the asexual life cycle show ring/dot-like structures (white arrow) within the FV, possibly degraded GFP-fusion proteins.Ehlgen F, Pham JS, de Koning-Ward T, Cowman AF, Ralph SA. Investigation of the Plasmodium falciparum food vacuole through inducible expression of the chloroquine resistance transporter (PfCRT). PLoS One. 2012;7(6):e38781.
See original on MMP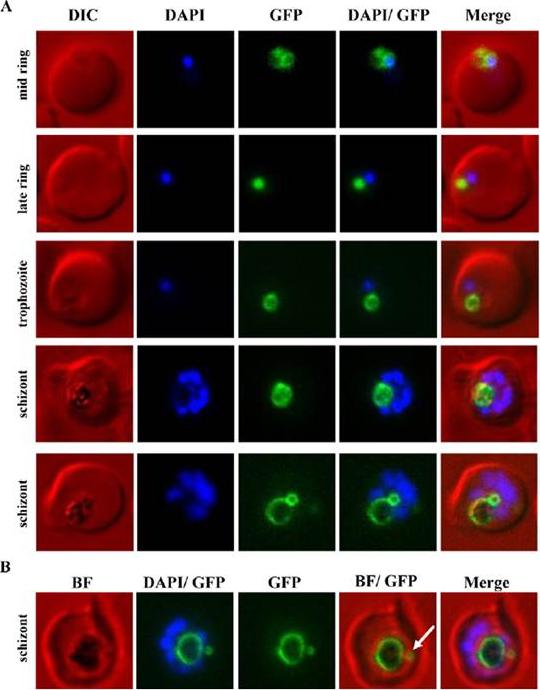
Analysis of the de novo biogenesis and development of the FV in live cells using PfCRT-GFP as a marker of the FV membrane. PfCRT was over-expressed as a GFP-fusion protein using an ATc-inducible expression system. (A) Live DAPI-stained cells were imaged. The earliest detectable and distinct localisation of PfCRT-GFP was observed in mid ring stage parasites, co-localising with a round spherical shape in proximity to the DAPI-stained nucleus. The characteristic dark haemozoin crystal was not yet visible in these parasites. In addition to FV labelling, some fluorescence was detectable at the ER. PfCRT-GFP labelling of the FV membrane was observed throughout the whole intra-erythrocytic life cycle. In accordance with an increase of the haemozoin crystal, the ring-like labelling of the FV membrane expanded as the parasite grew. An unexpected second PfCRT-GFP-labelled sphere was observed in schizont stage parasites. In some parasites this additional structure was associated/attached to the FV membrane (lower schizont panel). (B) An overlay with the corresponding brightfield (BF) image shows that in some parasites the additional PfCRT-GFP enclosed compartment (white arrow) is separate from the FV and surrounds a dark structure, possibly haemozoin.Ehlgen F, Pham JS, de Koning-Ward T, Cowman AF, Ralph SA. Investigation of the Plasmodium falciparum food vacuole through inducible expression of the chloroquine resistance transporter (PfCRT). PLoS One. 2012;7(6):e38781.
See original on MMP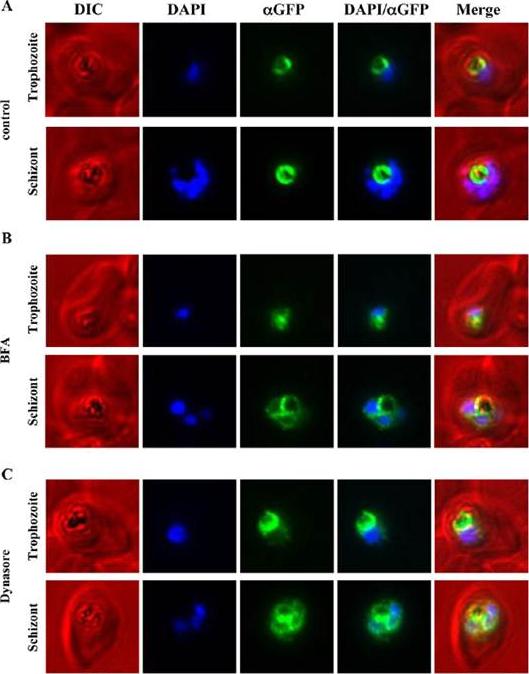
Immunofluorescence microscopy of PfCRT-GFP over-expressing parasites treated with either Brefeldin A or Dynasore. PfCRT was over-expressed as a GFP-fusion protein using an ATet-inducible expression system and treated with either BFA (5 mg/mL) for 3 h or Dynasore (40 mM/mL) for 2 h. As a control a second parasite population was treated with an equivalent volume of carrier alone (ethanol and DMSO, respectively). Following the BFA treatment, immunofluorescence microscopy was performed on fixed cells with mouse anti-GFP, Alexa-594 goat antimouse IgG and DAPI. Representative parasites in trophozoite and schizont stage are shown for control and treated parasites. (A) The control parasites show the restricted FV localisation of PfCRT-GFP (false-coloured in green). (B) BFA treated parasites show an accumulation of fluorescence around the DAPI-stained nuclei, consistent with ER localisation in addition to the FV localisation (Figure 1). (C) Treatment of parasites with Dynasore resulted in an accumulation of fluorescence around the DAPI-stained nuclei in addition to the FV membrane localisation, similar to the observed effect of BFA treatment (B).Ehlgen F, Pham JS, de Koning-Ward T, Cowman AF, Ralph SA. Investigation of the Plasmodium falciparum food vacuole through inducible expression of the chloroquine resistance transporter (PfCRT). PLoS One. 2012;7(6):e38781.
See original on MMP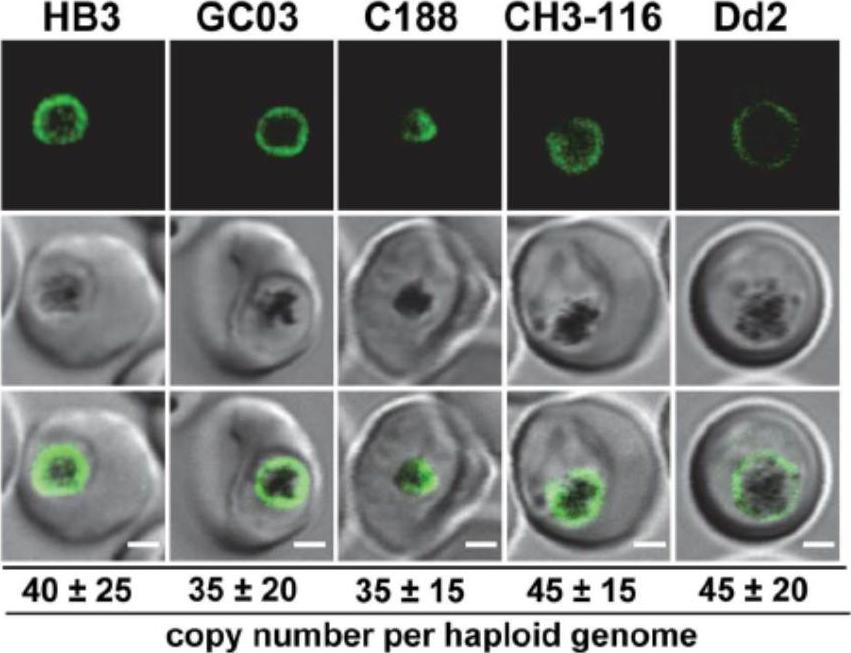
Functional association between the B5M12 locus and mutant PfCRT in conferring quinine and quinidine response variations. A. Live cell images of the P. falciparum strains indicated expressing an episomal copy of the Dd2 form of pfcrt fused to the GFP coding sequence. Fluorescence is located at the digestive vacuolar membrane, consistent with previous reports [69]. Bar, 2 mM. The copy number of the plasmid per haploid genome is indicated. The means 6 SEM of at least 6 biological replicates are shown. The copy numbers are not statistically different in the transfectants. we selected three progeny (GC03, CH3-116 and C188) that harbor the wild type pfcrt allele but which differ with regard to the B5M12 locus. GC03 contains the wild type HB3 B5M12 variant, whereas CH3-116 and C188 inherited the B5M12 locus from Dd2 (Table 1). The three progeny and the two parental clones HB3 and Dd2 were transfected with a vector expressing the Dd2 pfcrt variant fused in frame with the coding sequence of the green fluorescence protein. In all cases, the PfCRT/GFP fusion protein was expressed and localized at the membrane of the parasite’s digestive vacuole, as determined by live cell fluorescence microscopy.Sanchez CP, Liu CH, Mayer S, Nurhasanah A, Cyrklaff M, Mu J, Ferdig MT, Stein WD, Lanzer M. A HECT ubiquitin-protein ligase as a novel candidate gene for altered quinine and quinidine responses in Plasmodium falciparum. PLoS Genet. 2014 10(5):e1004382.
See original on MMP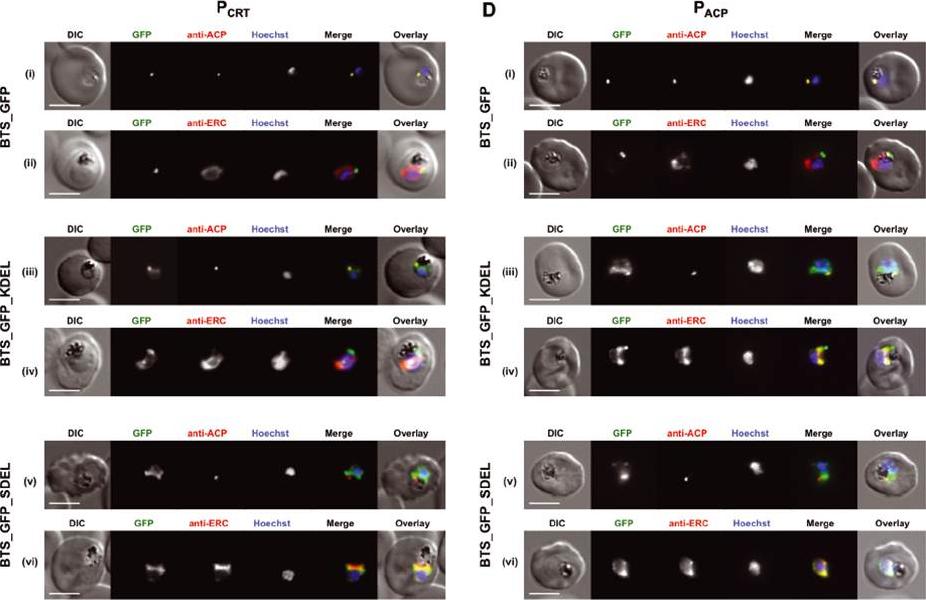
Addition of ER retrieval sequences to an apicoplast-targeted protein leads to reduced delivery to the apicoplast. A and C) Live-cell imaging of parasites expressing the apicoplast leader sequence of ACP (BTS) fused to GFP (top panel), GFP_KDEL (middle panel) and GFP_SDEL (both KDEL and SDEL are ER retention sequences; lower panel) under control of either the PfCRT (PCRT) or PfACP (PACP) promoter. B and D) Immunofluorescence assay of parasites expressing the apicoplast leader BTS fused to GFP (i, ii), GFP_KDEL (iii, iv) and GFP_SDEL (v, vi) as described above with anti-PfACP (apicoplast marker) and anti-PfERC (ER marker) antibodies. Scale bar: 3 μm (all).Heiny SR, Pautz S, Recker M, Przyborski JM. Protein Traffic to the Plasmodium falciparum Apicoplast: Evidence for a Sorting Branch Point at the Golgi. Traffic. 2014 15(12):1290-304
See original on MMP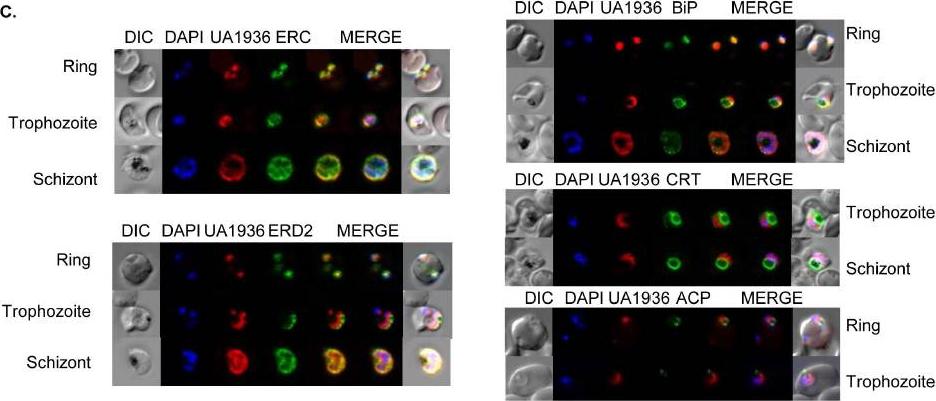
Fluorescence microscopy of UA1936(and inhibitor) and its localization in P. falciparum-infected red blood cells. P. falciparum-infected red blood cells (asynchronous cultures) were cultured with or without 100 mM UA1936 for 1 h and then fixed with 4% paraformaldehyde. In-cell click chemistry was performed with 2.5 mM Alexa 488 alkyne (green) or 2.5 TAMRA alkyne (red) for 30 min. For immunodetection, P. falciparum-infected red blood cells were incubated with specific antibodies against the endoplasmic reticulum markers ERC and BiP, against the cis-Golgi marker ERD2, against the food vacuole membrane marker CRT and against the apicoplast marker ACP.Penarete-Vargas DM, Boisson A, Urbach S, Chantelauze H, Peyrottes S, Fraisse L, Vial HJ. A chemical proteomics approach for the search of pharmacological targets of the antimalarial clinical candidate albitiazolium in Plasmodium falciparum using photocrosslinking and click chemistry. PLoS One. 2014 Dec 3;9(12):e113918. ·
See original on MMP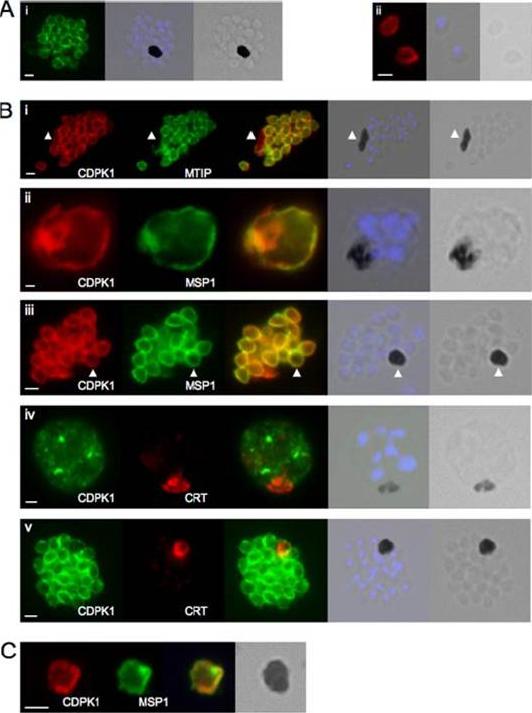
CDPK1 localizes to the plasma membrane of merozoites. A, an anti-CDPK1 antibody detects CDPK1 at the parasite periphery in schizonts (panel i) and free merozoites (panel ii). The second panel shows a merged image of nuclear staining with 4’,6’-diamino-2-phenylindole (blue) and the bright field image, and the third panel shows the bright field image. B, panel i, CDPK1 colocalizes with MTIP around the periphery of merozoites but is also found around the residual body, marked with an arrowhead, from which MTIP is absent. In young (panel ii) and segmented (panel iii) schizonts, CDPK1 colocalizes with the plasma membrane marker MSP1. Both proteins are also detected around the residual body, marked with an arrowhead. CDPK1 does not colocalize with the food vacuole marker PfCRT in either immature (panel iv) or mature (panel v) schizonts. The third panels in all of the images show the merged images of the antibody staining, beside which is a merged image of the bright field image and the nuclei of the parasites stained with 4’,6’-diamino-2-phenylindole (blue), followed by the bright field image alone. C, both CDPK1 (red) and MSP1 (green) can be detected on residual bodies that are released upon schizont rupture. The merged images of the antibody staining are shown in the third panels, followed by the bright field image. In all cases the white scale bar in the first panel of each set of images represents 1 mm.Green JL, Rees-Channer RR, Howell SA, Martin SR, Knuepfer E, Taylor HM, Grainger M, Holder AA. The motor complex of Plasmodium falciparum: phosphorylation by a calcium-dependent protein kinase. J Biol Chem. 2008 7;283(45):30980-9. PMID:
See original on MMP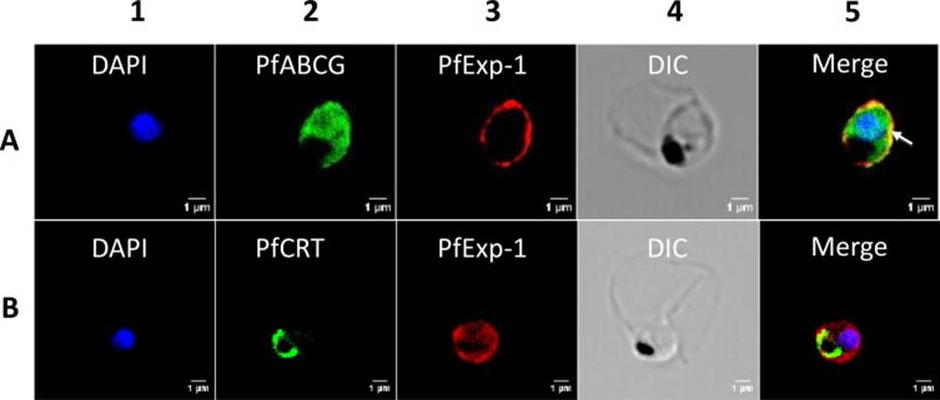
Co-localization of PfExp-1 and PfABCG in P. falciparum-infected erythrocytes. Double immuno-histochemical staining of infected erythrocytes at trophozoites stage with antibodies against PfABCG and PfExp-1 (Panel A) and PfCRT and PfExp-1 (Panel B). Column 1 shows the nuclear DNA staining with DAPI (blue). In columns 2 and 3, PfABCG signal (green) (A) and PfCRT signal (green) (B) are resolved with Alexa-fluor 488-labeled goat anti-rabbit secondary antibody, and PfExp-1 signal (red) (A and B) is resolved with Alexa-fluor 594-labeled goat anti-rabbit secondary antibody. Columns 4 and 5 show respectively the differential interference contrast (DIC) of infected erythrocyte and the merger of column 1, 2 and 3 without DIC. The digestive vacuole membrane staining of PfCRT does not overlap with PfExp-1 staining while the plasma membrane staining of PfABCG partially overlaps with the parasitophorous vacuole membrane signal of PfExp-1 as indicated by the arrow.Edaye S, Georges E. Characterization of native PfABCG protein in Plasmodium falciparum. Biochem Pharmacol. 2015 Jul 31. [Epub ahead of print] PMID:
See original on MMP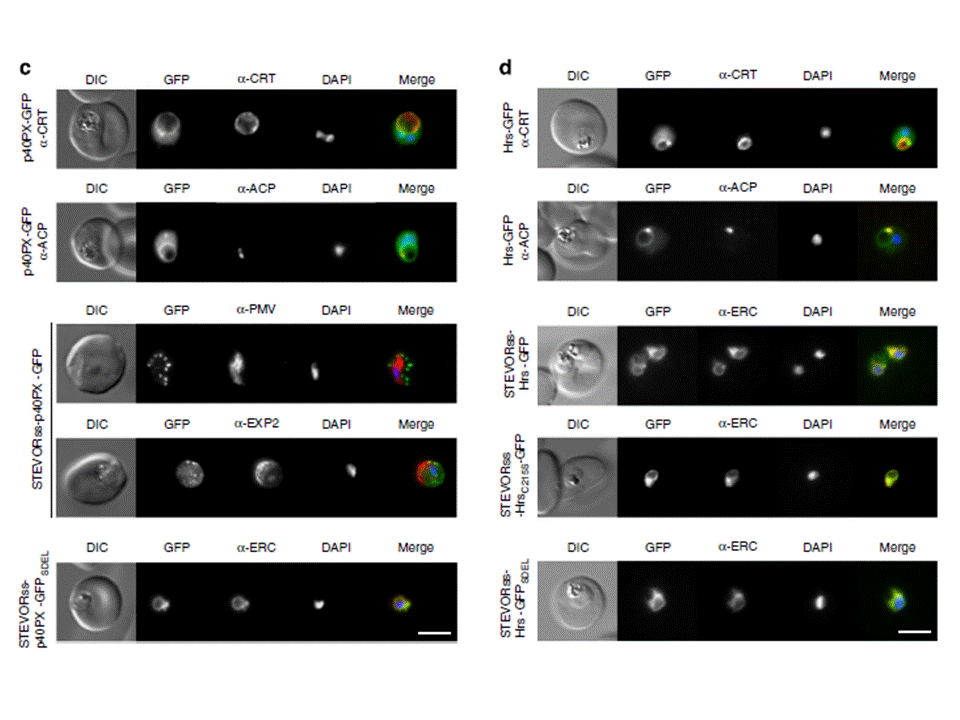
(c) p40PX-GFP localizes to the cytoplasm and to membranes of the food vacuole (labelled with anti-CRT) and apicoplast (labelled with anti-ACP). STEVORss-p40PX-GFP localizes to the PV in P. falciparum-infected erythrocytes, as shown by co-localization with anti-EXP2, and not in the ER, which was labelled with anti-PMV. If PI(3)P was located within the ER, this chimera would be expected to remain within the ER via p40PX binding. Fusion to an ER-retention signal (STEVORss-p40PX-GFPSDEL) retains the protein in ER, co-localization with anti-ERC. Scale bar, 5 mm.(d) Localization of Hrs-GFP to the cytoplasm and membranes of food vacuole (labelled anti-CRT) and apicoplast (labelled anti-ACP). STEVORss-Hrs-GFP localizes to the ER as shown by co-localization with anti-ERC; however, mutation of residues necessary for lipid binding in both Hrs FYVE fingers (STEVORss-HrsC215S-GFP) does not abrogate ER localization, demonstrating PI(3)P-independent retention in ER. Addition of ER retention signal (STEVORss-Hrs-GFPSDEL) also causes ER localization. Scale bar, 5 mm. Boddey JA, O'Neill MT, Lopaticki S, Carvalho TG, Hodder AN, Nebl T, Wawra S,van West P, Ebrahimzadeh Z, Richard D, Flemming S, Spielmann T, Przyborski J,Babon JJ, Cowman AF. Export of malaria proteins requires co-translationalprocessing of the PEXEL motif independent of phosphatidylinositol-3-phosphatebinding. Nat Commun. 2016 Feb 1;7:10470.
See original on MMPMore information
| PlasmoDB | PVP01_0109300 |
| GeneDB | PVP01_0109300 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | null |
| Orthologs | PBANKA_1219500 , PCHAS_1220200 , PF3D7_0709000 , PKNH_0107600 , PVX_087980 , PY17X_1222700 |
| Google Scholar | Search for all mentions of this gene |