PKNH_1221100 inner membrane complex sub-compartment protein 3, putative (ISP3)
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. berghei ANKA |
Possible |
RMgm-962 | Imported from RMgmDB | |
| P. yoelii yoelii 17X |
Possible |
RMgm-4890 | Imported from RMgmDB | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. berghei ANKA | Asexual |
No difference |
RMgm-962 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Asexual |
No difference |
RMgm-4890 | Imported from RMgmDB |
| P. berghei ANKA | Gametocyte |
No difference |
RMgm-962 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Gametocyte |
No difference |
RMgm-4890 | Imported from RMgmDB |
| P. berghei ANKA | Ookinete |
No difference |
RMgm-962 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Ookinete |
Difference from wild-type |
RMgm-4890
Both isp1 and isp3 mutants showed normal development of asexual blood stages and gametocytes in vivo as well as gamete formation and zygote-to-retort differentiation in vitro. However, the mature ookinete conversion in vitro was significantly reduced in isp1 (conversion rate: 34%) but not in isp3 (57%) compared with wild type (WT, 64%) |
Imported from RMgmDB |
| P. berghei ANKA | Oocyst |
No difference |
RMgm-962 | Imported from RMgmDB |
| P. berghei ANKA | Sporozoite |
No difference |
RMgm-962 | Imported from RMgmDB |
| P. berghei ANKA | Liver |
No difference |
RMgm-962 | Imported from RMgmDB |
Imaging data (from Malaria Metabolic Pathways)
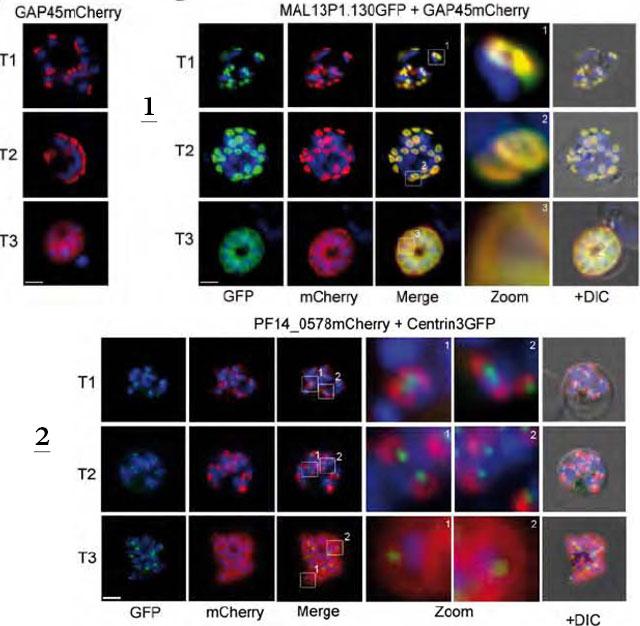
1. Co-localization of the TMD protein MAL13P1.130 and the glideosome associated protein GAP45. GAP45 was episomally expressed as mCherry fusion protein and localized in different schizont stages (left). Double transgenic cell line expressing MAL13P1.130-GFP and GAP45-mCherry reveals identical spatial distribution (right).2. Coexpression of the centrosome marker PfCentrin3-GFP and PF14_0578-mCherry shows a very close association between the forming IMC and the centrosome, as PfCentrin3 marks the centre of the developing IMC cramp-structure. Nuclei stained with DAPI. Scale bars, 2 μm.Kono M, Herrmann S, Loughran NB, Cabrera A, Engelberg K, Lehmann C, Sinha D, Prinz B, Ruch U, Heussler V, Spielman T, Parkinson J, Gilberger TW. Evolution and Architecture of the Inner Membrane Complex in Asexual and Sexual Stages of the Malaria Parasite. Mol Biol Evol. 2012 29(9):2113-32
See original on MMP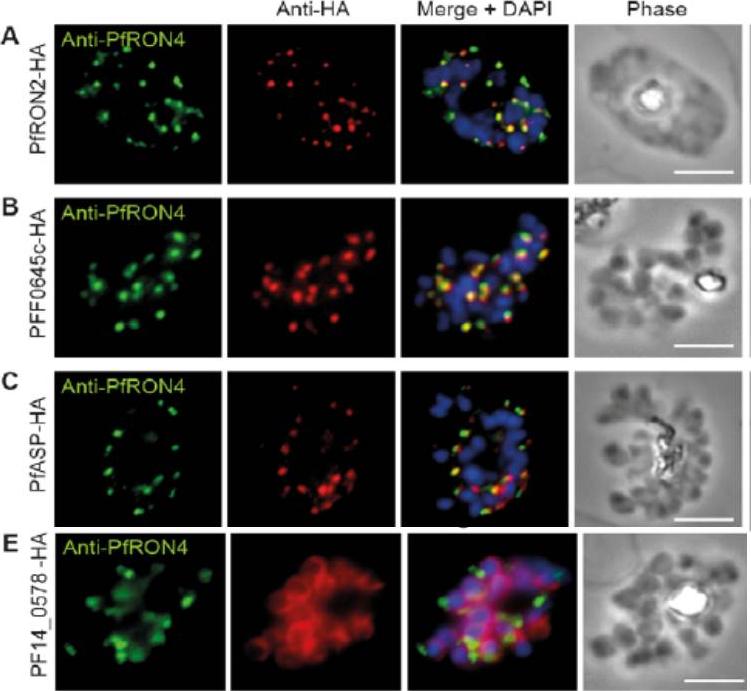
Cellular localisation of invasins in P. falciparum schizonts. (A–E) IFA of schizonts from HA tagged P. falciparum parasite lines labeled with anti-PfRON4, to mark the rhoptry neck, DAPI, to mark nuclei and anti-HA. (A) PfRON2 (B) rhoptry protein PFF0645c (C) apical sushi protein PfASP and (E) inner membrane complex protein PF14_0578. PfRON2-HA schizonts were colabeled with anti-PfRON4, a marker of the rhoptry neck. PFF0645c and PfASP each displayed a similar apical localization in schizonts, showing overlap of labeling with PfRON4. PF14_0578-HA labeling was peripheral in late stage schizonts (E), suggestive of an inner-membrane complex (IMC) or plasma membrane localization.Zuccala ES, Gout AM, Dekiwadia C, Marapana DS, Angrisano F, Turnbull L, Riglar DT, Rogers KL, Whitchurch CB, Ralph SA, Speed TP, Baum J. Subcompartmentalisation of Proteins in the Rhoptries Correlates with Ordered Events of Erythrocyte Invasion by the Blood Stage Malaria Parasite. PLoS One. 2012;7(9):e46160.
See original on MMP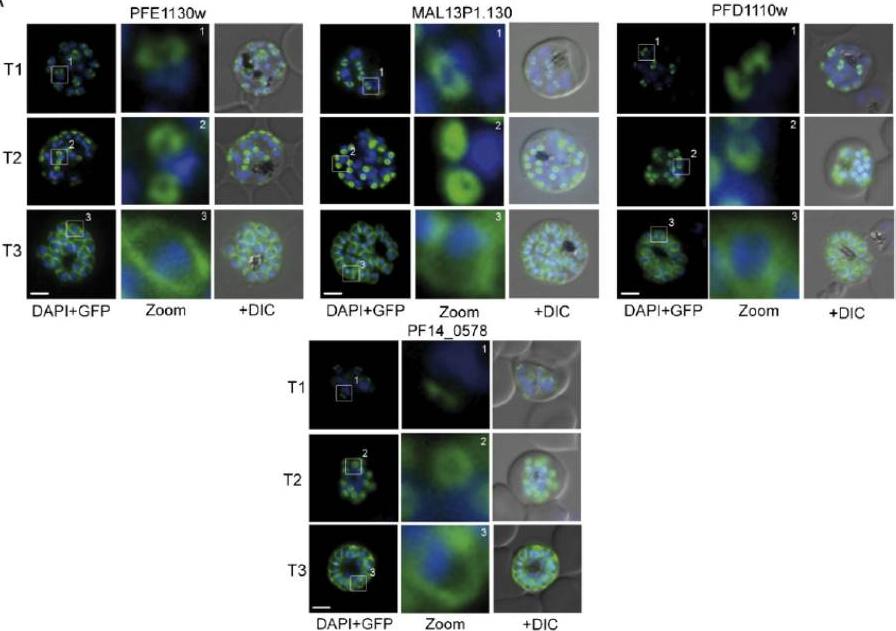
Several IMC proteins co-localize with GAP45 during schizogony. (A) Using GFP as a fluorescence tag, the TMD proteins PFE1130w, MAl13P1.130, PFD1110w, as well as PF14_0578 reveal three distinct structures during ongoing schizogony. Commencing as cramplike structures (T1), they transform to small ring-shaped formations (T2) that toward the end of schizogony expand and are then equally distributed underneath the plasma membrane (T3). Enlargement of these distinct structures are marked (white square) and shown in second row (Zoom) for each cell line.Kono M, Herrmann S, Loughran NB, Cabrera A, Engelberg K, Lehmann C, Sinha D, Prinz B, Ruch U, Heussler V, Spielmann T, Parkinson J, Gilberger TW. Evolution and architecture of the inner membrane complex in asexual and sexual stages of the malaria parasite. Mol Biol Evol. 2012 Sep;29(9):2113-32. PMID:
See original on MMP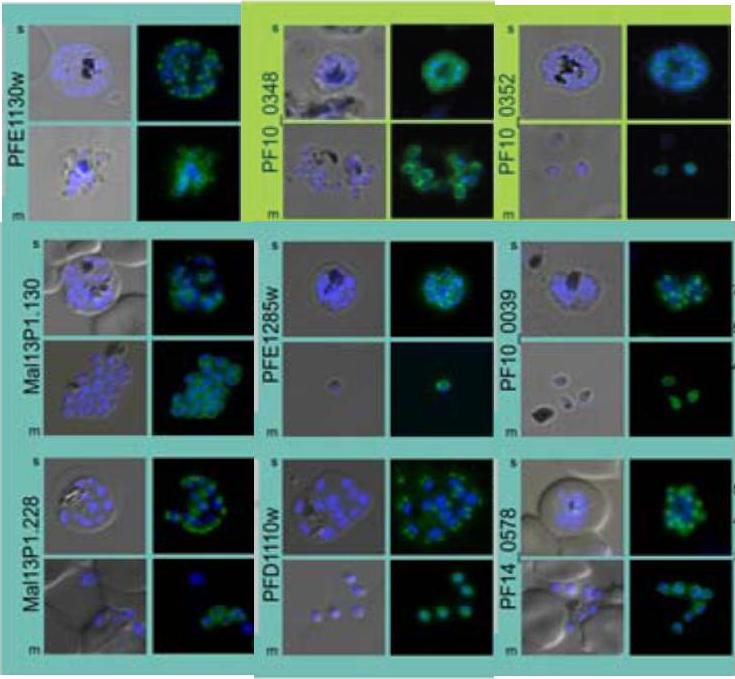
Subcellular distribution of proteins predicted to be involved in invasion. All proteins were localized in schizonts (s) and free merozoites (m) using GFP-fusion proteins and grouped according to their predominant GFP localization: surface (green), inner membrane complex (turquoise),Hu G, Cabrera A, Kono M, Mok S, Chaal BK, Haase S, Engelberg K, Cheemadan S, Spielmann T, Preiser PR, Gilberger TW, Bozdech Z. Transcriptional profiling of growth perturbations of the human malaria parasite Plasmodium falciparum. Nat Biotechnol. 2010 28(1):91-8.
See original on MMP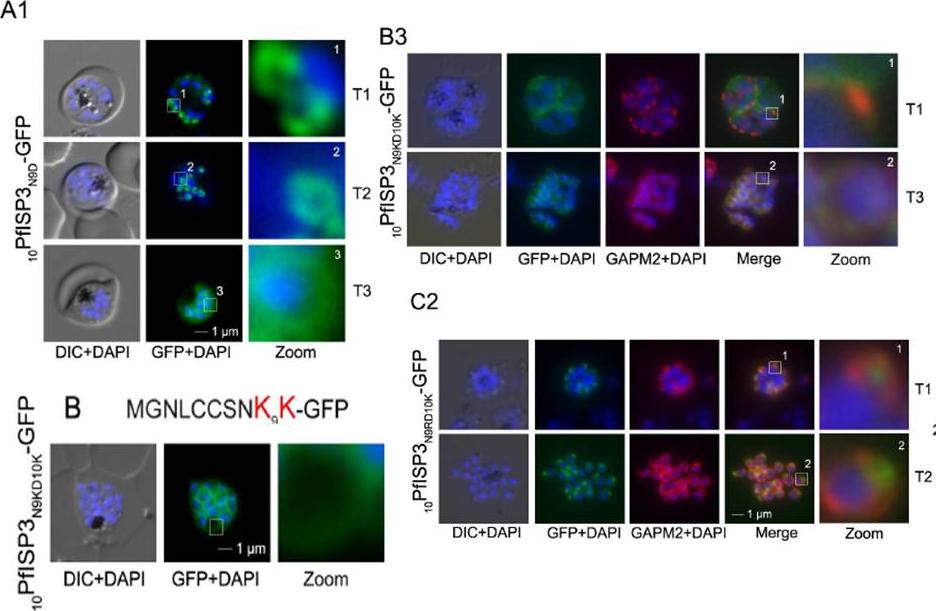
Mutational analysis of the minimal sequence requirements for IMC association of PfISP3. AB. Expression and localization of the 20 amino acid N-terminus with the last 10 (20PfISP311-20A-GFP) amino acids substituted with alanines. A1. 20PfISP311-20A-GFP was localized in unfixed parasites andshowed an identical dynamic as the wild type protein. A2. B. The first 10 amino acids of PFISP3 (10PfISP3-GFP) are sufficient for IMC localization (B1). C-D. Mutation of the sole charged residue D10 in to a neutral glycine (10PfISP3D10G-GFP, C1) or a positively charged arginine (10PfISP3D10R-GFP, D1) does not interfere with IMC targeting. Western blot analysis (C2, D2) of GFP fusion protein expression. Wetzel J, Herrmann S, Swapna LS, Prusty D, Peter AT, Kono M, Saini S, Nellimarla S, Wong TW, Wilcke L, Ramsay O, Cabrera A, Biller L, Heincke D, Mossman K, Spielmann T, Ungermann C, Parkinson J, Gilberger TW. The role of palmitoylation for protein recruitment to the inner membrane complex of the malaria parasite. J Biol Chem. 2014 Nov 25.
See original on MMP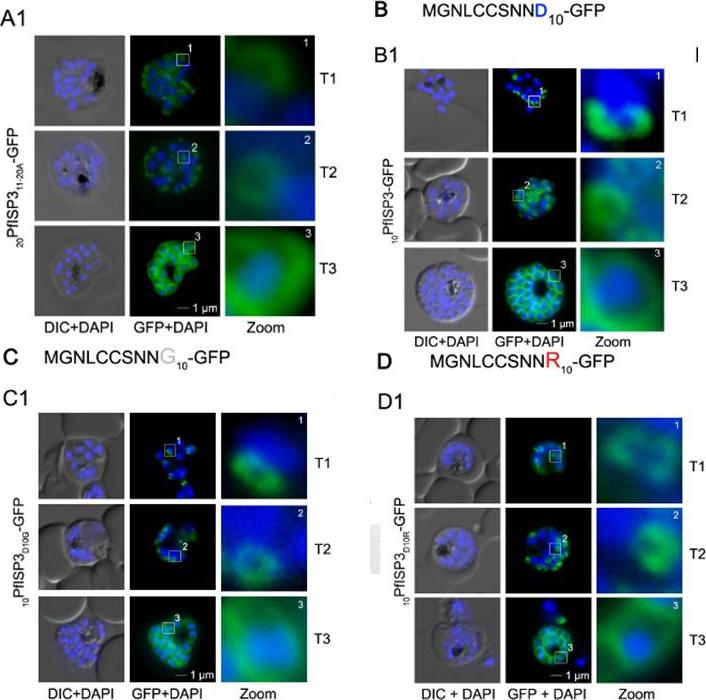
Mutational analysis of the minimal sequence requirements for IMC association of PfISP3. AB.Expression and localization of the 20 amino acid N-terminus with the last 10 (20PfISP311-20A-GFP) amino acids substituted with alanines. A1. 20PfISP311-20A-GFP was localized in unfixed parasites and showed an identical dynamic as the wild type protein.. B. The first 10 amino acids of PFISP3 (10PfISP3-GFP) are sufficient for IMC localization (B1) B2. Mutation of the sole charged residue D10 in to a neutral glycine (10PfISP3D10G-GFP, C1) or a positively charged arginine(10PfISP3D10R-GFP, D1) does not interfere with IMC targeting. Wetzel J, Herrmann S, Swapna LS, Prusty D, Peter AT, Kono M, Saini S, Nellimarla S, Wong TW, Wilcke L, Ramsay O, Cabrera A, Biller L, Heincke D, Mossman K, Spielmann T, Ungermann C, Parkinson J, Gilberger TW. The role of palmitoylation for protein recruitment to the inner membrane complex of the malaria parasite. J Biol Chem. 2014 Nov 25.
See original on MMP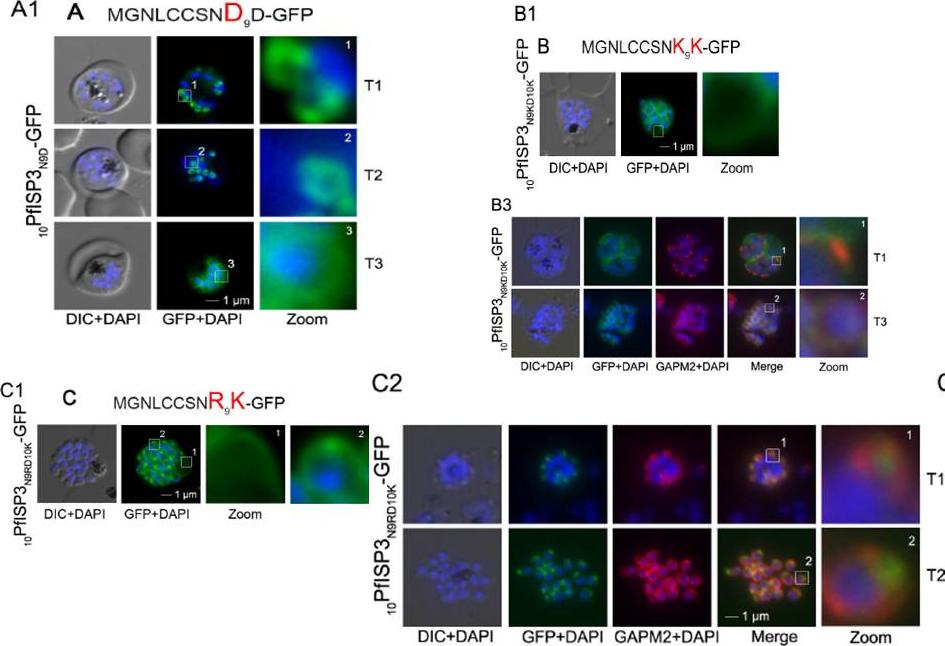
Re-direction of the IMC 10PfISP3-GFP fusion protein to the rhoptry membrane. A. Substitution of N9 with D (10PfISP3N9D-GFP, A) does not interfere with correct IMC recruitment of the GFP-fusion protein (A1). Nuclei stained with DAPI (blue). B-C. The substitution of N9 and D10 with lysine and its expression in the parasite (10PfISP3N9KD10K-GFP) leads to a peripheral localization (B1). The zoom highlights the additional association of this mutant protein with the food vacuole membrane. B2. Western blot analysis using anti-GFP antibodies. B3 Co-localization with the IMC marker GAPM2. Nascent IMC (anti-GAPM2, red) is clearly distinguishable from the peripheral associated 11PfISP1D11K-GFP in early stages (T1) and congruent in late stages (T3). C. In contrast, the substitution of N9 with arginine (10PfISP3N9RD10K-GFP) re-directs the fusion protein to cell periphery (zoom 1) and the apical pole (zoom 2). C2. Co-localization with the rhoptry marker RALP (anti-RALP, red) in fixed cells confirmed PfDHHC1-GFP IMC localization shown here in an early stage of TMC biogenesis (T1). Wetzel J, Herrmann S, Swapna LS, Prusty D, Peter AT, Kono M, Saini S, Nellimarla S, Wong TW, Wilcke L, Ramsay O, Cabrera A, Biller L, Heincke D, Mossman K, Spielmann T, Ungermann C, Parkinson J, Gilberger TW. The role of palmitoylation for protein recruitment to the inner membrane complex of the malaria parasite. J Biol Chem. 2014 Nov 25. PMID:
See original on MMP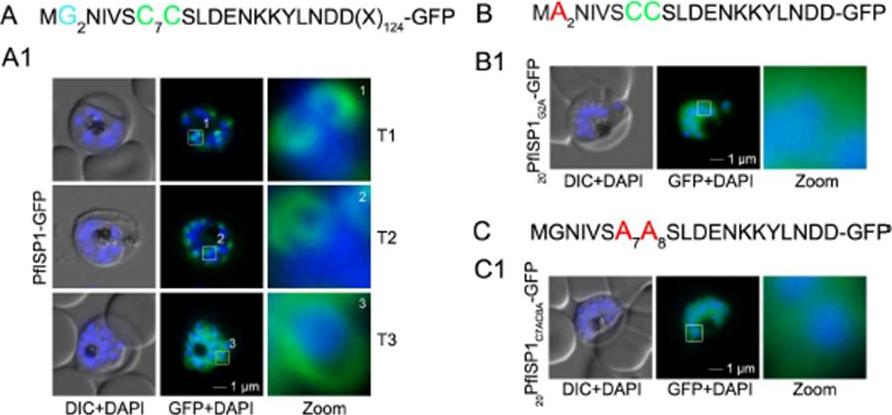
The role of N-terminal acylation for IMC membrane association of PfISP1 and acyl biotin exchange assays. A. Over-expression of PfISP1-GFP showing the same characteristic IMC dynamics as PfISP3-GFP. Putative myristoylation and palmitoylation sites are highlighted in light blue (G2) or green (C7C8). B-C. Mutation of either one of the acylation motifs (20PfISP1G2A-GFP and 20PfISP1C7AC8A-GFP) resulted in a cytosolic variant shown by microscopy. Nuclei are stained with DAPI (blue). Enlargement of selected areas are marked with a white square and referred to as zoom. Scale bar, 1 μm. Wetzel J, Herrmann S, Swapna LS, Prusty D, Peter AT, Kono M, Saini S, Nellimarla S, Wong TW, Wilcke L, Ramsay O, Cabrera A, Biller L, Heincke D, Mossman K, Spielmann T, Ungermann C, Parkinson J, Gilberger TW. The role of palmitoylation for protein recruitment to the inner membrane complex of the malaria parasite. J Biol Chem. 2014 Nov 25.
See original on MMP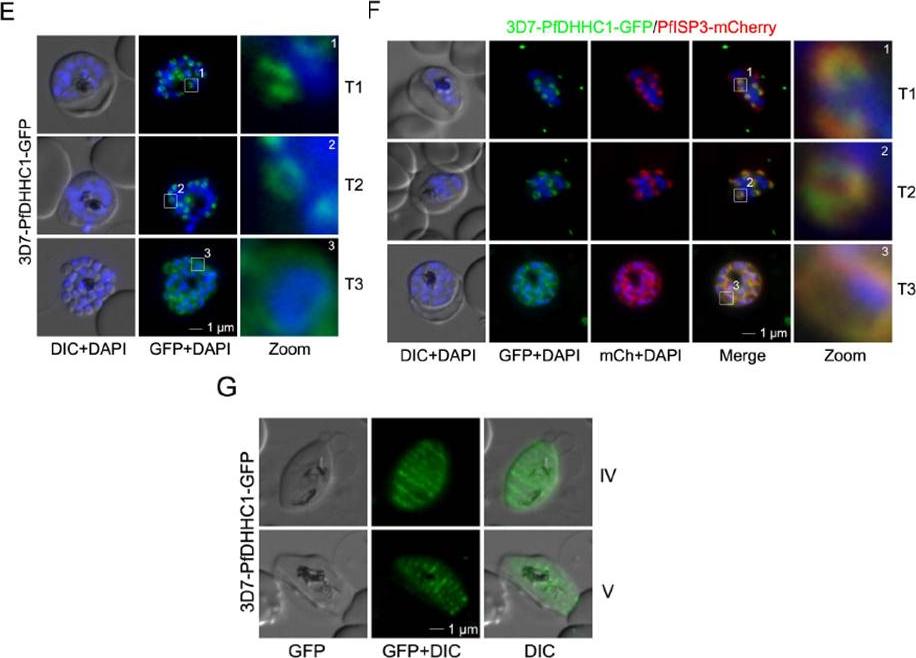
PfDHHC1 is an IMC localized palmitoyl acyltransferase. E. PfDHHC1-GFP was localized in unfixed late stage parasites showing characteristic IMC dynamics during schizogony (T1-T3). Nuclei stained with DAPI (blue). Enlargement of selected areas are marked with a white square and referred as zoom. Scale bar, 1 μm. F. Co-localization of PfDHHC1-GFP and PfISP3mCherry. PfISP3mCherry was episomally co-expressed and shows identical spatial distribution compared with PfDHHC1-GFP. G. PfDHHC1-GFP distribution within the nascent IMC during gametocytog-enesis (stage IV and V). These symmetric sutures of the IMC are expanding with ongoing growth of the IMC vesicles and maturation of the gametocytes (stage III and IV). Scale bars, 1 μm.Wetzel J, Herrmann S, Swapna LS, Prusty D, Peter AT, Kono M, Saini S, Nellimarla S, Wong TW, Wilcke L, Ramsay O, Cabrera A, Biller L, Heincke D, Mossman K, Spielmann T, Ungermann C, Parkinson J, Gilberger TW. The role of palmitoylation for protein recruitment to the inner membrane complex of the malaria parasite. J Biol Chem. 2014 Nov 25. PMID:
See original on MMP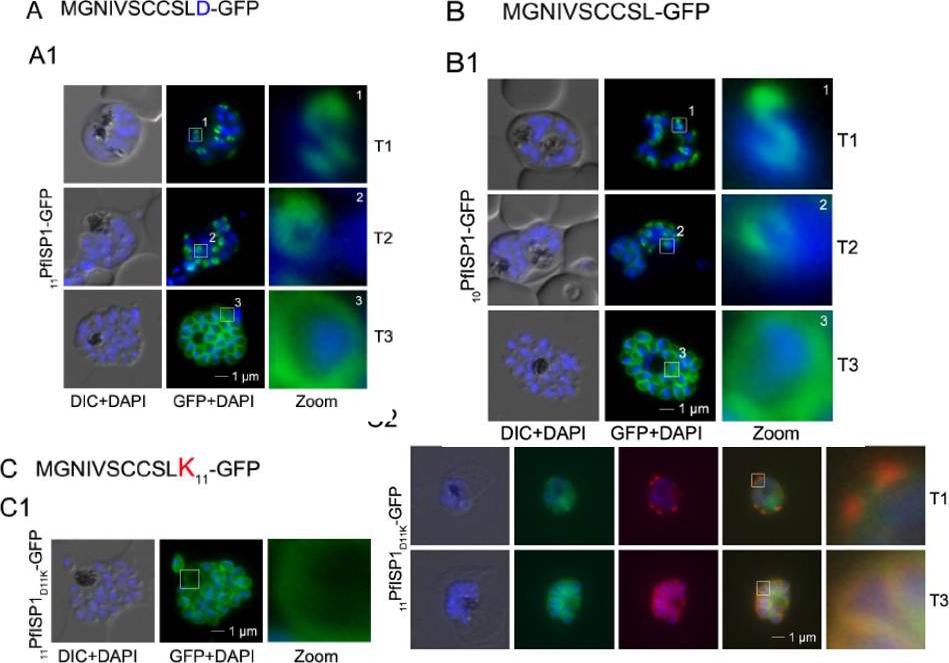
Mutational analysis of the minimal sequence requirements for IMC association of PfISP1. AB. Expression of the first 11 (11PfSP1-GFP, A) or 10 (10PfISP1-GFP, B) amino acids are sufficient to direct GFP to the IMC (A1-B1). C. Substitutionof D11 with lysine (11PfISP1D11K-GFP, C) abolishes IMC targeting (C1) and leads to peripheral localization. The zoom highlights its additional association with the food vacuole membrane consistent with plasma membrane association. C2. Co-localization with the IMC marker GAPM2. Nascent IMC (anti-GAPM2, red) is clearly distinguishable from the peripheral associated 11PfISP1D11K-GFP in early stages (T1) and congruent in late stages (T3). Wetzel J, Herrmann S, Swapna LS, Prusty D, Peter AT, Kono M, Saini S, Nellimarla S, Wong TW, Wilcke L, Ramsay O, Cabrera A, Biller L, Heincke D, Mossman K, Spielmann T, Ungermann C, Parkinson J, Gilberger TW. The role of palmitoylation for protein recruitment to the inner membrane complex of the malaria parasite. J Biol Chem. 2014 Nov 25.
See original on MMP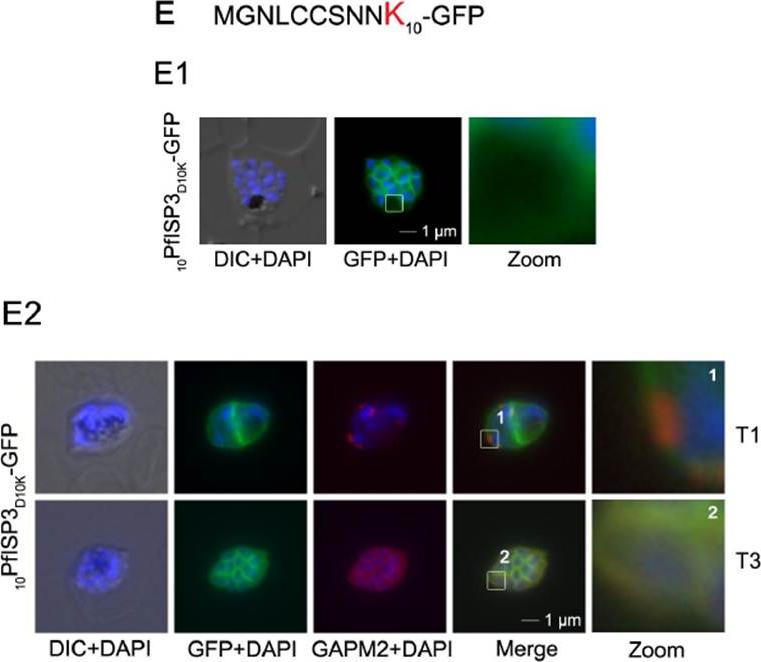
Mutational analysis of the minimal sequence requirements for IMC association of PfISP3. E. Substitution of D10 with a positively charged lysine re-directs the GFP-fusion protein to the periphery of the nascent merozoites. The zoom highlights the additional association with the food vacuole membrane characteristic for plasma membrane proteins (E1). Co-localization with the IMC marker GAPM2. Nascent IMC (anti-GAPM2, red) is clearly distinguishable from the peripheral associated 10PfISP3D10K-GFP in early stages (T1) and congruent in late stages (T3). Wetzel J, Herrmann S, Swapna LS, Prusty D, Peter AT, Kono M, Saini S, Nellimarla S, Wong TW, Wilcke L, Ramsay O, Cabrera A, Biller L, Heincke D, Mossman K, Spielmann T, Ungermann C, Parkinson J, Gilberger TW. The role of palmitoylation for protein recruitment to the inner membrane complex of the malaria parasite. J Biol Chem. 2014 Nov 25. PMID:
See original on MMP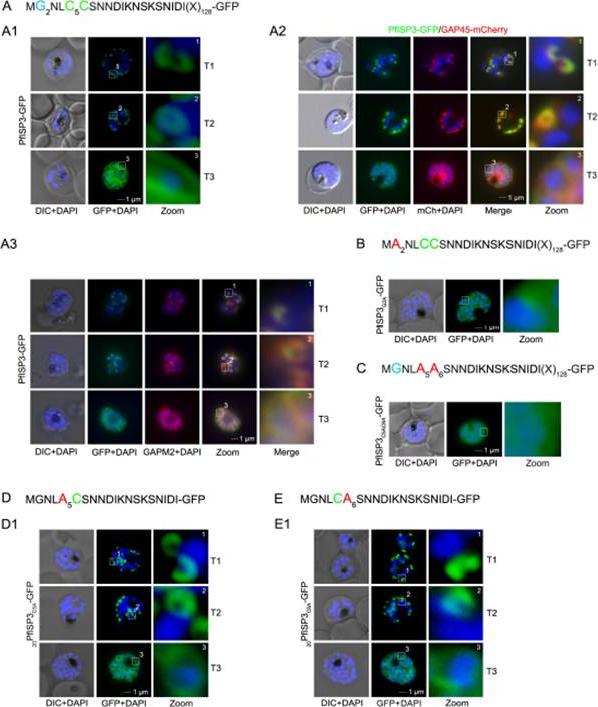
The role of N-terminal acylation for IMC membrane association of PfISP3. A. Over-expression of PfISP3-GFP showing characteristic IMC dynamics during schizogony (T1-T3) in unfixed parasites. Nuclei are stained with DAPI (blue). Enlargement of selected areas are marked with a white square and referred to as zoom. Scale bar, 1 μm (A1). A2. Co-localization of PfISP3-GFP (green) and GAP45mCherry (red) in unfixed parasites revealed their identical dynamic during schizogony. A3. Colocalization with the IMC marker GAPM2 (α-GAPM2, red) in fixed cells. Putative myristoylation and palmitoylation sites are highlighted in light blue (G2) or green (C5C6). B-C. IMC membrane association depends on the presence of N-terminal myristoylation and palmitoylation motifs. N-terminal myristoylation (B, PfISP3G2A-GFP) and palmitoylation motif mutants (C, PfISP3C5AC6A-GFP) were expressed in P. falciparum and localized in unfixed parasites. Mutation of either the myristoylation or the palmitoylation motifs resulted in a cytosolic distribution of the GFP-fusion protein (B1, C1). D-E. Mutation of the individual palmitoylation sites C5 (PfISP3C5A-GFP) or C6 (PfISP3C6A-GFP) does not interfere with IMC recruitment of the fusion protein. Wetzel J, Herrmann S, Swapna LS, Prusty D, Peter AT, Kono M, Saini S, Nellimarla S, Wong TW, Wilcke L, Ramsay O, Cabrera A, Biller L, Heincke D, Mossman K, Spielmann T, Ungermann C, Parkinson J, Gilberger TW. The role of palmitoylation for protein recruitment to the inner membrane complex of the malaria parasite. J Biol Chem. 2014 Nov 25.
See original on MMPMore information
| PlasmoDB | PKNH_1221100 |
| GeneDB | PKNH_1221100 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | PK14_2755w, PKH_124540 |
| Orthologs | PBANKA_1324300 , PCHAS_1327600 , PF3D7_1460600 , PVP01_1246200 , PVX_117425 , PY17X_1328100 |
| Google Scholar | Search for all mentions of this gene |