PKNH_0906200 endoplasmic reticulum-resident calcium binding protein, putative (ERC)
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. falciparum 3D7 |
Refractory |
USF piggyBac screen (Insert. mut.) | USF PiggyBac Screen | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. falciparum 3D7 | Asexual |
Egress defect |
32098818 (Knock down)
'Knockdown of PfERC shows that this gene is essential for asexual growth of P. falciparum. Analysis of the intraerythocytic lifecycle revealed that PfERC is required for parasite egress and invasion. We found that PfERC knockdown prevents the rupture of the parasitophorous vacuole membrane. This is because PfERC knockdown inhibited the proteolytic maturation of the subtilisin-like serine protease, SUB1, and the essential SUB1 substrate, the merozoite surface protein 1. PfERC knockdown further inhibited the proteolytic maturation of the essential invasion ligand, Apical Membrane Antigen 1 (AMA1), which occurs during egress.' |
Theo Sanderson, Google AI |
| P. falciparum 3D7 | Asexual |
Egress defect |
https://www.biorxiv.org/content/early/2018/11/01/457481
(Knock down)
PVM fails to rupture |
Theo Sanderson, Google AI |
Imaging data (from Malaria Metabolic Pathways)
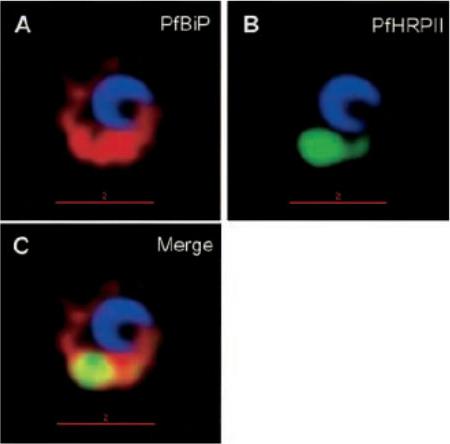
Protein accumulation in the ER in P. falciparum-infected cells treated with brefeldin A. Infected red cells at early times of asexual growth, were incubated with 5 mgml-1 brefeldin A for 24 h and subsequently stained for PfHRPII (green), PfBiP (red), Hoechst (blue for nuclei), in indirect immunofluorescence assays and imaged by DeltaVision deconvolution microscopy. Yellow indicates extent of merge between green and red. The location to the ER may reflect altered (albeit reversible) membrane organization induced by the drug, rather than a specialized organelle. In addition, since the P. falciparum (primary) ER is not well characterized, the context of a secondary ER remains unclear.Haldar K, Mohandas N, Samuel BU, Harrison T, Hiller NL, Akompong T, Cheresh P. Protein and lipid trafficking induced in erythrocytes infected by malaria parasites. Cell Microbiol. 2002 4:383-95.
See original on MMP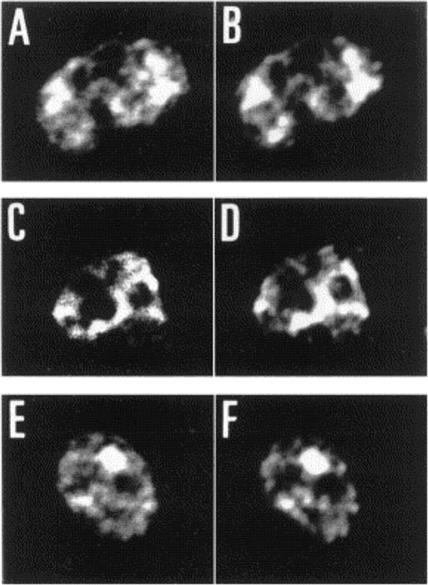
Confocal immunofluorescence co-localization of PfERC with the ER-located protein, Pfgrp (PFI0875w). Mature trophozoite-infected erythrocytes (D10 strain, Panels A–D or FCR3strain, Panels E–F) were double-labeled with mouse antiserum against recombinant Pfs40:PfERC produced in either yeast (Panels A,E) or E. coli (Panel C), followed by FITC-conjugated anti-mouse antibodies and a rabbit anti-Pfgrp antiserum followed by a Cy3-conjugated anti-rabbit antibodies (Panels B,D,F). An optical slice was collected through the centre of the parasites by confocal microscopy. Dual images were collected simultaneously and are displayed as isolated fluorescein fluorescence (Panels A,C,E) and isolated Cy3 fluorescence (Panels B,D,F) of the same cell. Labeling pattern suggests localization of PfERC within structures within the parasite cytoplasm. Staining profile for the known ER-resident protein, Pfgrp exactly co-localize with PfERC.La Greca N, Hibbs AR, Riffkin C, Foley M, Tilley L. Identification of an endoplasmic reticulum-resident calcium-binding protein with multiple EF-hand motifs in asexual stages of Plasmodium falciparum. Mol Biochem Parasitol. 1997 89:283-93. Copyright Elsevier
See original on MMP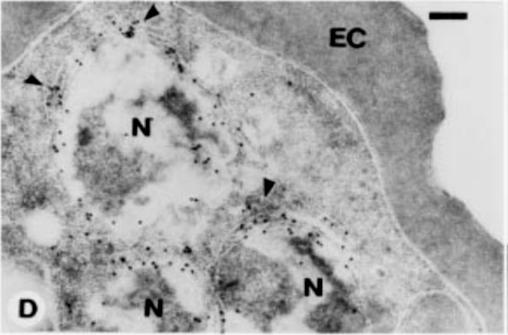
Immunoelectron microscopy of LR White sections of asexual schizont stage were fixed in 1% paraformaldehyde:0.2% glutaraldehyde and incubated with sera (M40) against E. coli-produced Pfs40 MBP-fusion protein. Arrows point to labeling with gold particles. Nucleus (N); erythrocyte cytoplasm (EC). Bar equals 2.0 mM. This result indicates localization in a membrane-bound compartment adjacent to the nucleus that is suggestive of endoplasmic reticulumTempleton TJ, Fujioka H, Aikawa M, Parker KC, Kaslow DC. Plasmodium falciparum Pfs40, renamed Pf39, is localized to an intracellular membrane-bound compartment and is not sexual stage-specific. Mol Biochem Parasitol. 1997 ;90:359-65. Copyright Elsevier 2009.
See original on MMP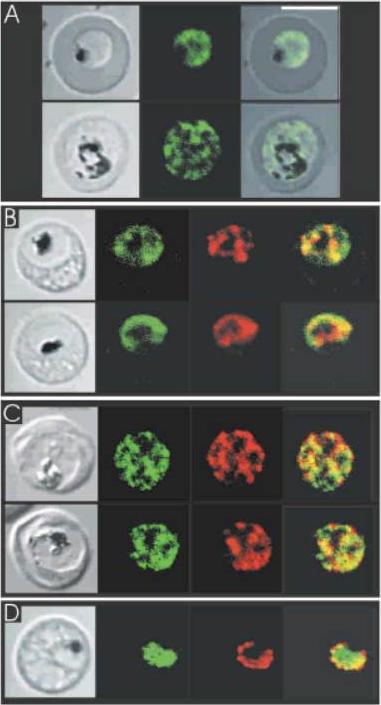
Intracellular location of Pfd-COP and comparison with the locations of the ER-located protein, PfERC, the trans -Golgi marker, PfRab6, and the PV-located protein, Exp-1. (A) Erythrocytes infected with young trophozoite stage (top panels) or mature trophozoite stage (bottom panels) malaria parasites (FAC8 strain) were labeled with affinity purified rabbit anti-GST-Pfd-COP(90) antiserum followed by an FITC-conjugated anti-rabbit IgG. (B) Erythrocytes infected with trophozoite stage parasites (FAC8 strain) were labeled with affinity purified rabbit anti-GST-Pfd-COP(90) antiserum followed by an FITC-conjugated anti-rabbit IgG (green fluorescence) and a murine antiserum recognizing the ER-located protein, PfERC, followed by Alexa Fluor 568-conjugated anti-mouse IgG (red fluorescence). (C) Erythrocytes infected with trophozoite stage parasites (FAC8 strain) were labeled with affinity purified rabbit anti-GST-Pfd-COP(90) antiserum followed by an FITC-conjugated anti-rabbit IgG (green fluorescence) and a murine MAb recognizing the Golgi protein, PfRab6, followed by Alexa Fluor 568-conjugated anti-mouse IgG (red fluorescence). (D) Erythrocytes infected with trophozoite stage parasites (FAC8 strain) were labeled with affinity purified rabbit anti-GST-Pfd-COP(90) antiserum followed by an FITC-conjugated anti rabbit IgG (green fluorescence) and a murine MAb recognizing the PV located protein, Exp-1 PF11_0224, followed by Alexa Fluor 568-conjugated antimouse IgG (red fluorescence). Differential interference contrast (DIC) images are shown in the left hand panels. Overlays of the DIC and green fluorescence channels are shown in the right hand panels of (A). Overlays of the green and red fluorescence channels are shown in the right hand panels of (B-D). Optical slices were collected by confocal microscopy through the centre of the parasite. The bar in the top right hand panel represents 5 mm.Adisa A, Rug M, Foley M, Tilley L. Characterisation of a delta-COP homologue in the malaria parasite, Plasmodium falciparum. Mol Biochem Parasitol. 2002 123:11-21. Copyright Elsevier 2010.
See original on MMP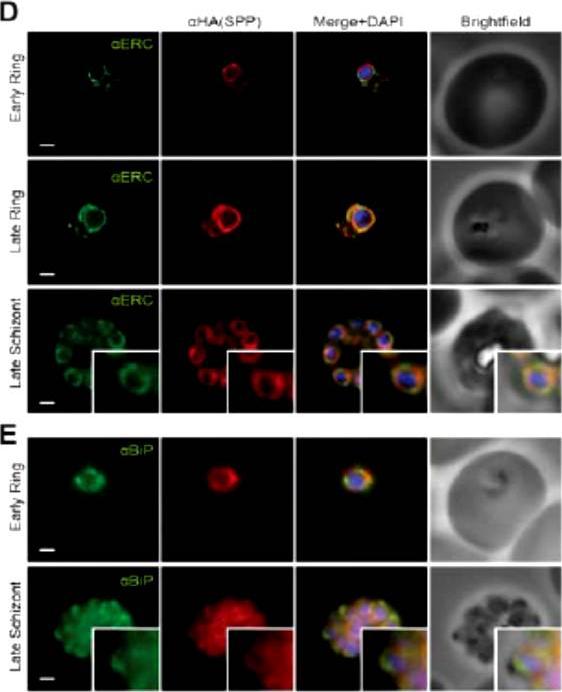
PfSPP localises to the parasite nuclear periphery. IFA of ring, trophozite and schizont stages of PfSPP-HA labeled with anti-PfERC and HA antibodies. Scale bar 1 μm. Co-labeling with ER markers BiP and ER-calcium binding protein (ERC) by IFA showed strong coincidental labeling with PfSPP fluorescence across the entire intra-erythrocytic development of the parasite. SPP is localized to the ER.Marapana DS, Wilson DW, Zuccala ES, Dekiwadia CD, Beeson JG, Ralph SA, Baum J. Malaria parasite signal peptide peptidase is an ER-resident protease required for growth but not invasion. Traffic. 2012 13(11):1457-65
See original on MMP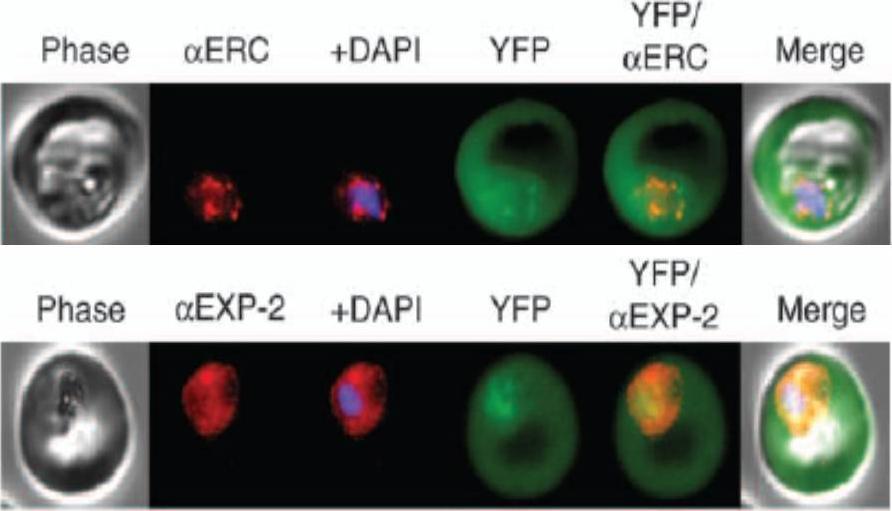
KAHRP chimaeras localise to the endoplasmic reticulum in addition to the parasitophorous vacuole or erythrocyte cytosol. Images captured by immunofluorescence microscopy show substantial colocalisation of KAHRP chimaeras with the endoplasmic reticulum calcium-binding protein ERC (upper panel). Intraparasitic fluorescence is also evident when chimaeras were colocalised with the parasitophorous vacuole membrane protein EXP-2 PF14_0678 (lower panel). Only the wild type (WT) PEXEL chimaera efficiently exported to the erythrocyte cytosol,Boddey JA, Moritz RL, Simpson RJ, Cowman AF. Role of the Plasmodium export element in trafficking parasite proteins to the infected erythrocyte. Traffic. 2009 10:285-99. Copyright John Wiley & Sons Ltd. 2010.
See original on MMP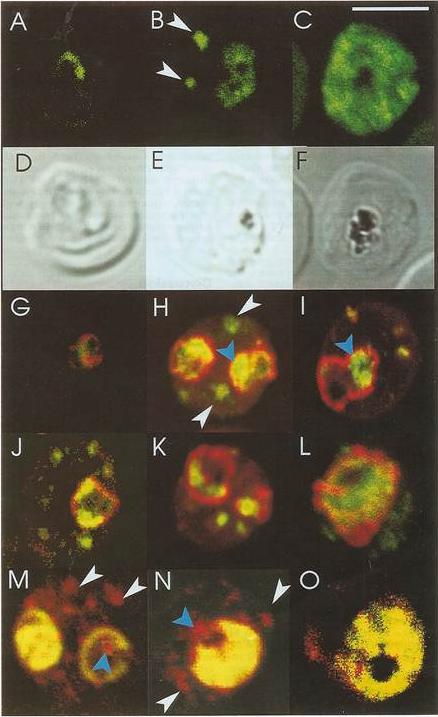
Confocal immunofluorescence of PfSar1p. Panels A-C: Parasitized erythrocyes (D10 strain) were labeled with affinity purified anti=PfSar1p antiserum followed by fluorescein-conjugated anti-rabbit antiserum (green). Panels D-F: Transmission images of panels A-C. Panels G-L: Dual labeling of affinity-purified rabbit anti-PfSar1p antiserum followed by fluorescein-conjugated anti-rabbit antiserum (green) and a murine monoclonal antibody recognizing the parasitophorous vacuole-located protein, Exp-1, followed by rhodamine-conjugated anti-mouse antiserum (red). Sar1p in the trophozoite is clearly located in the erythrocyte cytosol (G-K) and in the schizont it occupies most of it (L). Panels M-O: Dual labeling of affinity-purified rabbit anti-PfSar1p antiserum followed by rhodamine-conjugated anti-mouse antiserum (red) and a biotin-conjugated anti-PfERC antiserum followed by fluorescein-conjugated anti-rabbit antiserum (green). PfERC and PfSar1p labeling patterns do not completely coincide, indicating that PfSar1p is not confined to the ER. An optical slice was collected through the center of the parasite by confocal microscopy. Some PfSar1p-containing structures in the erythrocyte cytosol are marked with white arrowheads. Some PfSar1p-containing structures in the parasite cytosol are marked with blue arrowheads. The bar in panel C corresponds to 5 mm.Albano FR, Berman A, La Greca N, Hibbs AR, Wickham M, Foley M, Tilley L. A homologue of Sar1p localises to a novel trafficking pathway in malaria-infected erythrocytes. Eur J Cell Biol. 1999 78:453-462. Copyright Elsevier 2010.
See original on MMP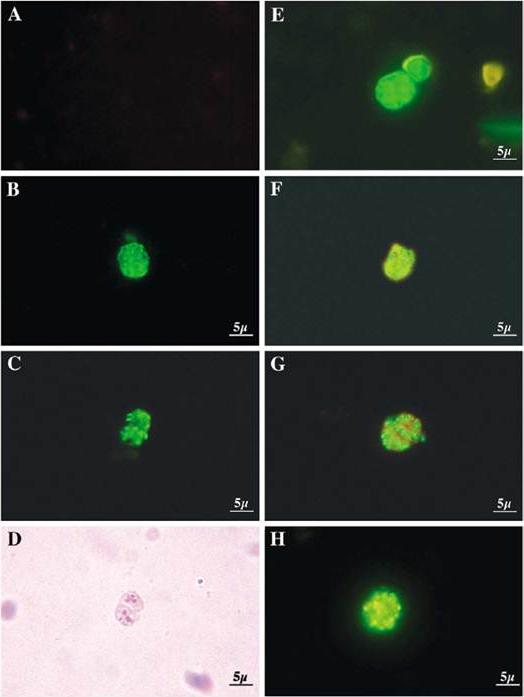
Different PfRabs have different sub-cellular distributions in schizont-infected erythrocytes. Panel (A) shows control rabbit pre-immune serum. Panel (B) shows a multinucleated schizont decorated with anti-PfRab1A antibodies giving a perinuclear staining. Panels (C) and (D) (phase contrast) show the pattern obtained with the anti-PfRab11A antibodies. Clearly, PfRab1A and PfRab11A display different distributions. Panel (E) shows the pattern obtained with anti-Pf39 (ERC) antibodies specific for the endoplasmic reticulum. Panel (F) shows a double-labelling with anti-PfRab1A (green) and Pf39 (red) antibodies, where colocalization stains yellow. Panel (G) shows a double-labelling with anti-PfRab11A (green) and anti-Pf39 (red) and no co-localization is observed. Panel (H) shows anti-PfRab6 (green) and anti-Pf39 (red), where co-localization stains yellow.Quevillon E, Spielmann T, Brahimi K, Chattopadhyay D, Yeramian E, Langsley G. The Plasmodium falciparum family of Rab GTPases. Gene. 2003 306:13-25.
See original on MMP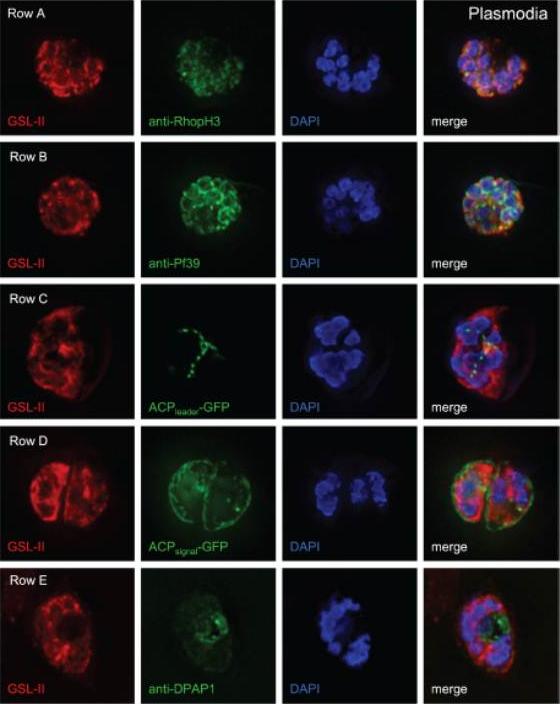
Deconvolving micrographs of Plasmodium-infected RBCs show that GSL-II, which binds to Plasmodium N-glycans, colocalizes with rhoptries (row A) and the ER (row B) but does not colocalize with apicoplasts (row C), the parasitophorous vacuole (row D), or the food vacuole (row E). In each case GSL-II is labeled red with Alexafluor 594, while antibodies labeled green with Alexafluor 488 are against RhopH3 (rhoptries), Pf39 (ER), and food vacuole (DPAP1). Alternatively, GFP is targeted to apicoplasts (ACPleader-GFP) and the parasitophorous vacuole (ACPsignal-GFP). Control experiments with anti-GFP antibodies show that both apicoplasts and parasitophorous vacuoles are accessible to exogenous probes. The nature of the glycosylated proteins is not knownBushkin GG, Ratner DM, Cui J, Banerjee S, Duraisingh MT, Jennings CV, Dvorin JD, Gubbels MJ, Robertson SD, Steffen M, O'Keefe BR, Robbins PW, Samuelson J. Suggestive evidence for Darwinian Selection against asparagine-linked glycans of Plasmodium falciparum and Toxoplasma gondii. Eukaryot Cell. 2010 9:228-41.
See original on MMP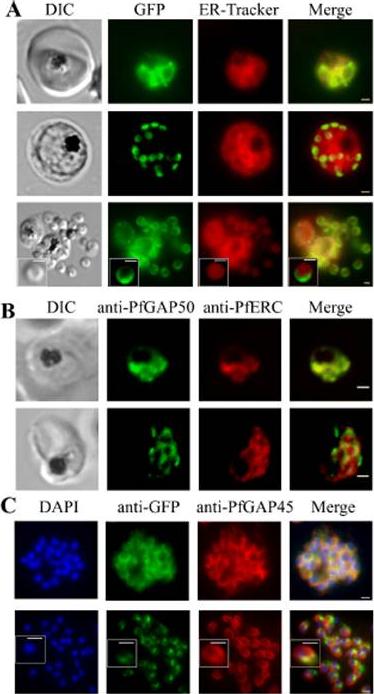
PfGAP50 is located in the ER prior to recruitment to the IMC. A. Parasites expressing PfGAP50-GFP (green) were incubated with ER-Tracker (depicted in red). The GFP and ER-Tracker fluorescence is co-located in trophozoite stage parasites. Upon relocation of GFP fluorescence to the apical ends of nascent merozoites during early schizont stage, the reticular ER-Tracker labelling persists. PfGAP50-GFP is concentrated at one pole of released merozoites while ER Tracker labels internal structures. B. Fixed cell immunofluorescence microscopy of the 3D7 parent strain labelled with antibodies raised against PfGAP50 (green) and PfERC (red). C. Immuno-fluorescence microscopy of PfGAP50-GFP transfectants at the mature and ruptured schizont stages labelled with anti-GFP (green), anti-PfGAP45 (red) and DAPI (blue). Bars = 1 μm. PfGAP50-GFP is located in the endoplasmic reticulum in early stage parasites, then redistributes to apical caps during the formation of daughter merozoites. In the final stage of schizogony the PfGAP50-GFP profile extends further around the merozoite surfaceYeoman JA, Hanssen E, Maier AG, Klonis N, Maco B, Baum J, Turnbull L, Whitchurch CB, Dixonand MW, Tilley L. Tracking glideosome-associated protein-50 reveals the development and organization of the inner membrane complex of P. falciparum. Eukaryot Cell. 2011 10(4):556-64.
See original on MMP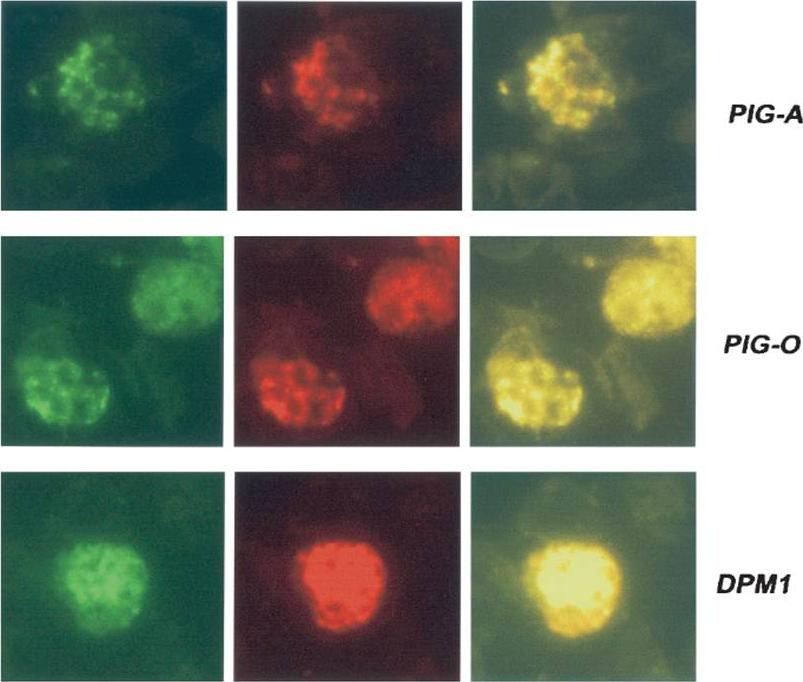
Detection of proteins in blood stage parasites was performed using indirect fluorescent-antibody tests. Thin films of parasites at mature stages were fixed in cold acetone and incubated sequentially with 1/80 dilutions of sera from peptide-immunized mice plus rabbit anti-ERC1 (1:200) to determine localization to the parasite ER. These slides were washed and incubated with an appropriate mixture of fluorescein-conjugated anti-mouse and rhodamine-conjugated anti-rabbit (1:1.000) antibodies. Slides were photographed under appropriate illumination for fluorescein isothiocyanate (first column, showing PIG-A, PIG-O, or DPM1 localization) and rhodamine (second column, showing ERC1 localization), and photographs were overlaid (third column).Delorenzi M, Sexton A, Shams-Eldin H, Schwarz RT, Speed T, Schofield L. Genes for glycosylphosphatidylinositol toxin biosynthesis in Plasmodium falciparum. Infect Immun. 2002 70:4510-22. PMID:
See original on MMP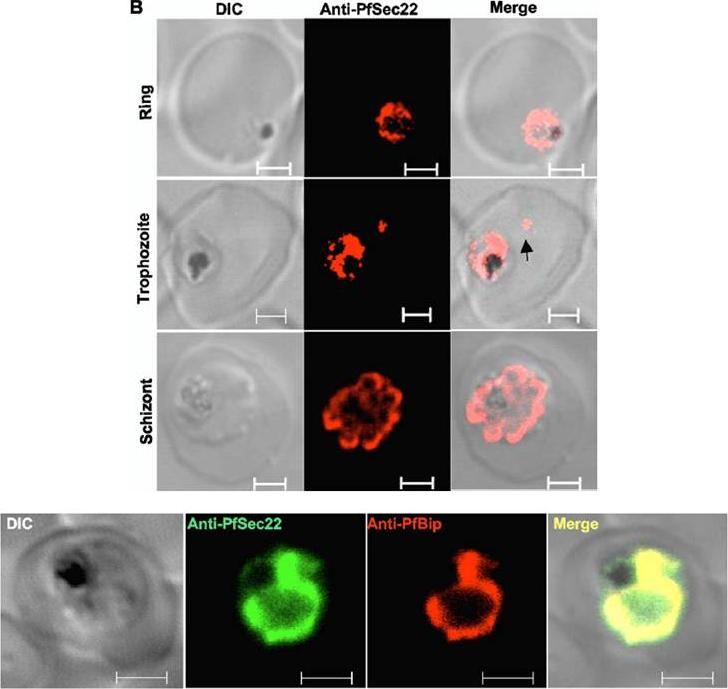
Immunofluorescence microscopy of fixed cells using anti-PfSec22 antibodies and goat anti-rabbit Alexa Fluor 555 secondary antibodies. An intense ring of PfSec22 fluorescence is visible in ring- and trophozoite-infected cells. Isolated foci of PfSec22 fluorescence (arrow) are also detected in the host cell compartment in trophozoite-infected cells, suggesting export of PfSec22 into the erythrocyte cytosol. Scale bars, 2 mm.Bottom row: Coimmunolocalization and membrane association of endogenous PfSec22. The association of PfSec22 with the parasite ER was investigated by coimmuno-fluorescence assays using rabbit anti-PfSec22 and rat anti-PfBip antibodies, as indicated. Both proteins colocalized significantly within the parasite. Scale bars, 2 mm.Ayong L, Raghavan A, Schneider TG, Taraschi TF, Fidock DA, Chakrabarti D. The Longin domain regulates the steady-state dynamics of Sec22 in P. falciparum. Eukaryot Cell. 2009 8(9):1330-40.
See original on MMP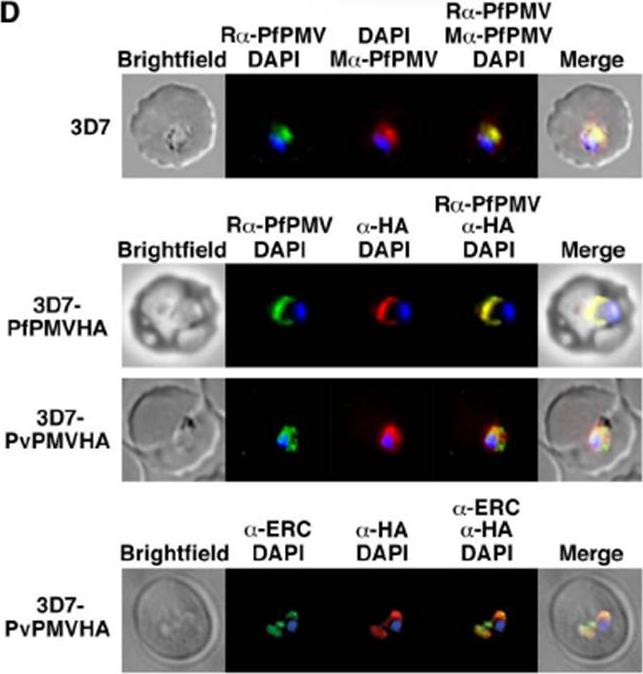
(D, Top) Immunofluorescence micrographs show rabbit a-PfPMV antibodies (Ra-PfPMV, green) label PfPMV in the ER. Colocalizations were performed with mouse a-PfPMV antibodies (Ma-PfPMV, red), shown previously to label PMV in the ER . (Middle) a-HA antibodies (red) label PfPMVHA (Top) and PvPMVHA (Bottom). P. vivax) in the parasite ER. (Bottom) a-HA antibodies (red) label PvPMVHA in the ER, as shown by clocalization with ERC (green). A strong perinuclear signal was observed in parasites expressing PvPMVHA or PfPMVHA (middle panels, red). Both proteins colocalized with rabbit anti-PfPMV antibodies, indicating the location was the ER. PvPMVHA also colocalized with ERC (bottom), as shown previously for PfPMVHA.Sleebs BE, Lopaticki S, Marapana DS, O'Neill MT, Rajasekaran P, Gazdik M, Günther S, Whitehead LW, Lowes KN, Barfod L, Hviid L, Shaw PJ, Hodder AN, Smith BJ, Cowman AF, Boddey JA. Inhibition of Plasmepsin V Activity Demonstrates Its Essential Role in Protein Export, PfEMP1 Display, and Survival of Malaria Parasites. PLoS Biol. 2014 12(7):e1001897.
See original on MMP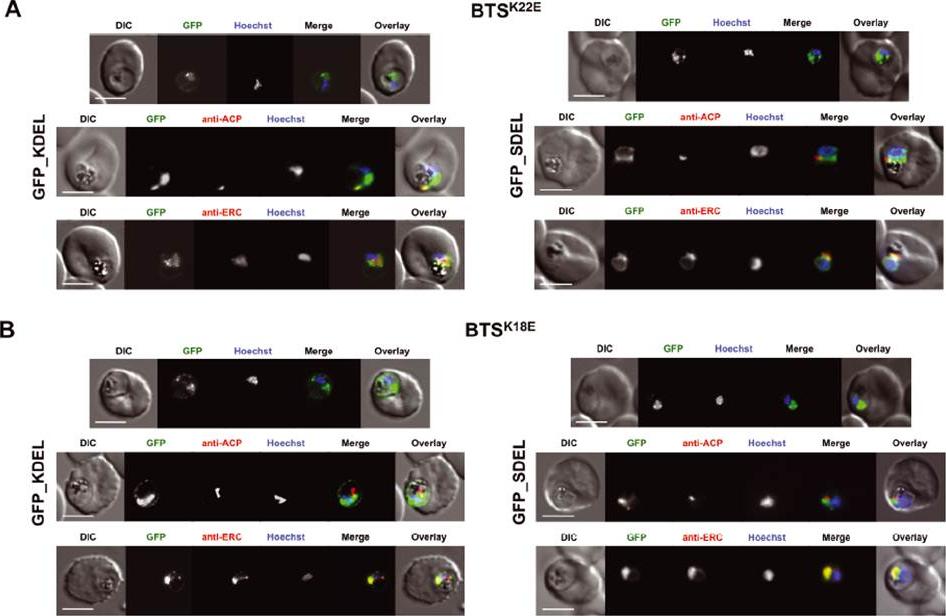
Analysis of ER retrieval strength compared to mutant transit peptides. A) Imaging of parasites expressing the K22E mutant form of ACP apicoplast leader sequence (BTSK22E) fused to GFP_KDEL (left panel) or GFP_SDEL (right panel). Upper panel: live-cell imaging; middle and lower panels: immunofluorescence assay using anti-PfACP or anti-PfERC. B) Imaging of parasites expressing the K18E mutant form of ACP apicoplast leader sequence (BTSK18E) fused to GFP_KDEL (left panel) or GFP_SDEL (right panel). Upper panel: live-cell imaging; middle and lower panels: immunofluorescence assay using anti-PfACP or anti-PfERC. Scale bar: 3 μm.Heiny SR, Pautz S, Recker M, Przyborski JM. Protein Traffic to the Plasmodium falciparum Apicoplast: Evidence for a Sorting Branch Point at the Golgi. Traffic. 2014 5(12):1290-304.
See original on MMP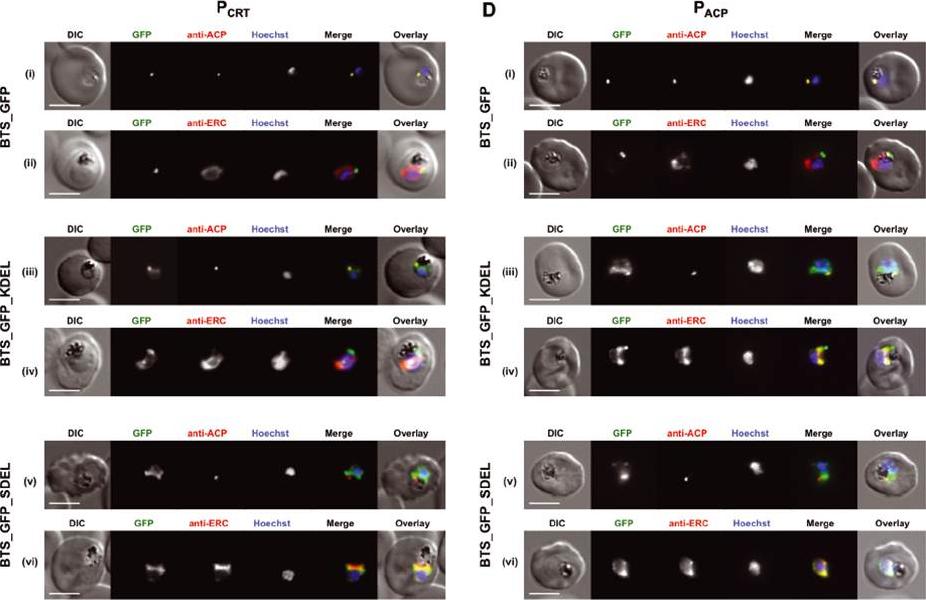
Addition of ER retrieval sequences to an apicoplast-targeted protein leads to reduced delivery to the apicoplast. A and C) Live-cell imaging of parasites expressing the apicoplast leader sequence of ACP (BTS) fused to GFP (top panel), GFP_KDEL (middle panel) and GFP_SDEL (both KDEL and SDEL are ER retention sequences; lower panel) under control of either the PfCRT (PCRT) or PfACP (PACP) promoter. B and D) Immunofluorescence assay of parasites expressing the apicoplast leader BTS fused to GFP (i, ii), GFP_KDEL (iii, iv) and GFP_SDEL (v, vi) as described above with anti-PfACP (apicoplast marker) and anti-PfERC (ER marker) antibodies. Scale bar: 3 μm (all).Heiny SR, Pautz S, Recker M, Przyborski JM. Protein Traffic to the Plasmodium falciparum Apicoplast: Evidence for a Sorting Branch Point at the Golgi. Traffic. 2014 15(12):1290-304
See original on MMP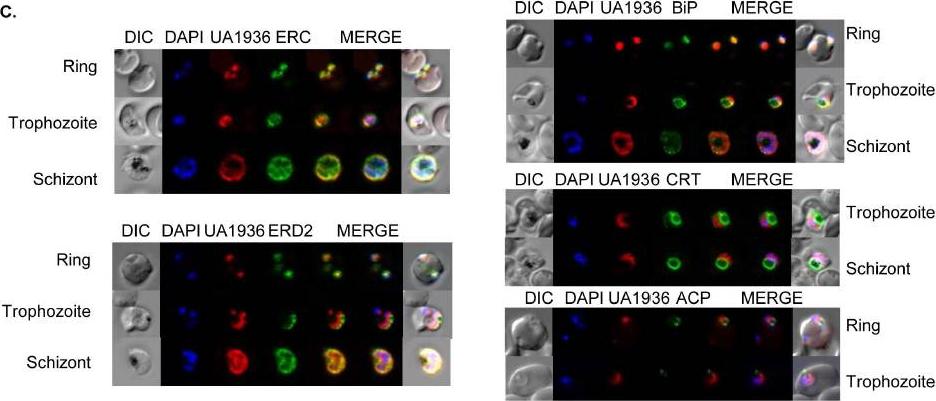
Fluorescence microscopy of UA1936(and inhibitor) and its localization in P. falciparum-infected red blood cells. P. falciparum-infected red blood cells (asynchronous cultures) were cultured with or without 100 mM UA1936 for 1 h and then fixed with 4% paraformaldehyde. In-cell click chemistry was performed with 2.5 mM Alexa 488 alkyne (green) or 2.5 TAMRA alkyne (red) for 30 min. For immunodetection, P. falciparum-infected red blood cells were incubated with specific antibodies against the endoplasmic reticulum markers ERC and BiP, against the cis-Golgi marker ERD2, against the food vacuole membrane marker CRT and against the apicoplast marker ACP.Penarete-Vargas DM, Boisson A, Urbach S, Chantelauze H, Peyrottes S, Fraisse L, Vial HJ. A chemical proteomics approach for the search of pharmacological targets of the antimalarial clinical candidate albitiazolium in Plasmodium falciparum using photocrosslinking and click chemistry. PLoS One. 2014 Dec 3;9(12):e113918. ·
See original on MMP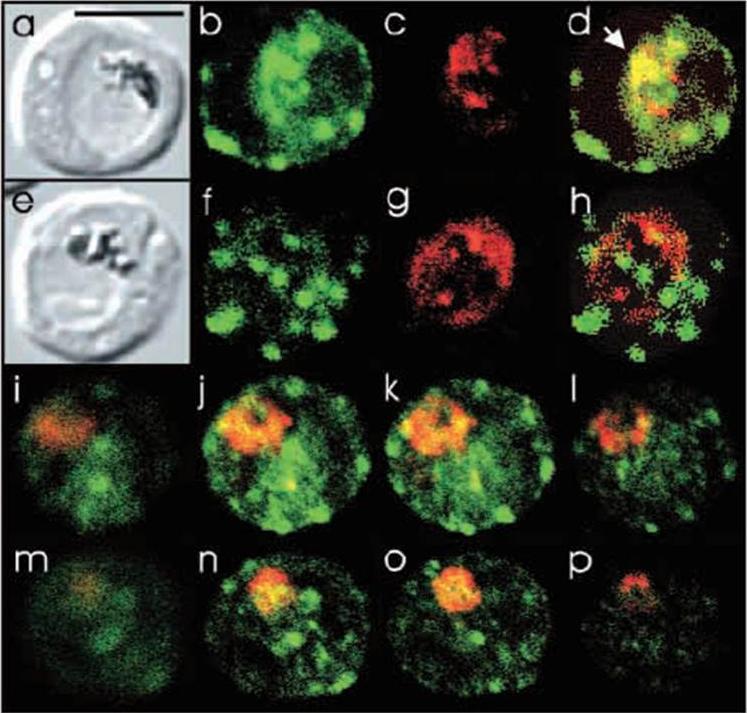
Comparison of the location of PfSec31p with the ER-located protein PfERC in P.-falciparum-infected erythrocytes and semiquantitative analysis of the amount of PfSec31p in the erythrocyte cytosol. (a-h) Asynchronous infected erythrocytes (K1 strain) were probed with affinity-purified rabbit anti-PfSec31(WD) antiserum followed by a fluorescein-conjugated anti-rabbit IgG (green fluorescence) and a murine antiserum recognising the ER located protein PfERC followed by Alexa-Fluor-568-conjugated antimouse IgG (red fluorescence). A region of partial overlap ofPfSec31p and PfERC in the parasite cytosol in (d) is marked with an arrowhead. (i-p) Asynchronous infected erythrocytes (K1 strain) were probed with rabbit anti-PfSec31(WD) antiserum (green fluorescence; i-l) or a murine anti-PfERC (red fluorescence; m-p). Four optical slices (0.6 mm) were collected by confocal microscopy from the bottom to the top of the parasitised erythrocytes.Adisa A, Albano FR, Reeder J, Foley M, Tilley L. Evidence for a role for a Plasmodium falciparum homologue of Sec31p in the export of proteins to the surface of malaria parasite-infected erythrocytes. J Cell Sci. 2001 114(Pt 18):3377-86.
See original on MMP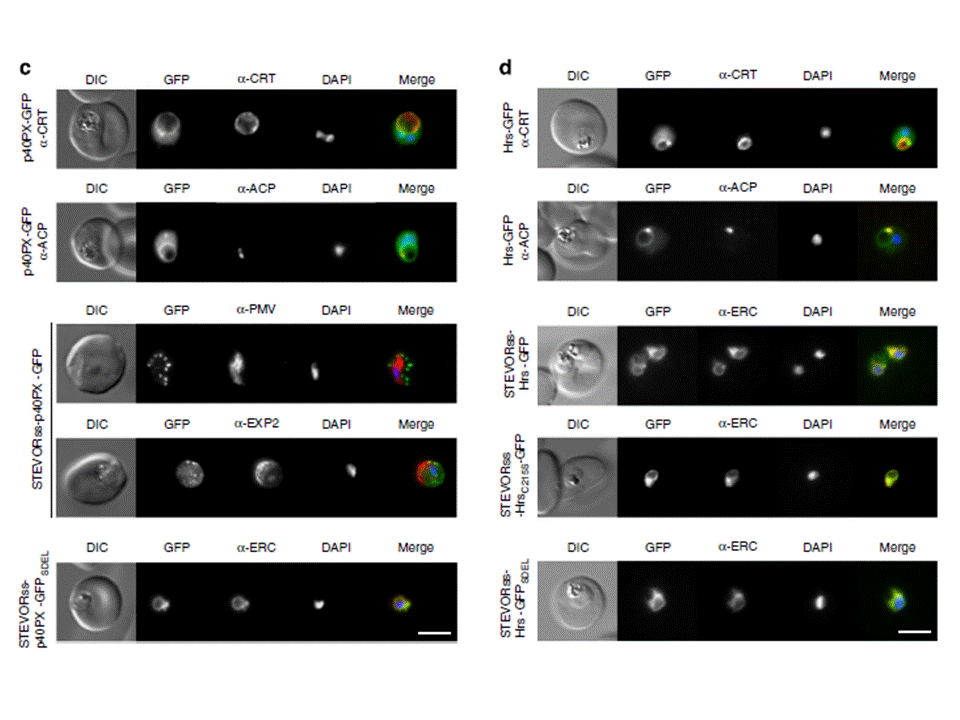
(c) p40PX-GFP localizes to the cytoplasm and to membranes of the food vacuole (labelled with anti-CRT) and apicoplast (labelled with anti-ACP). STEVORss-p40PX-GFP localizes to the PV in P. falciparum-infected erythrocytes, as shown by co-localization with anti-EXP2, and not in the ER, which was labelled with anti-PMV. If PI(3)P was located within the ER, this chimera would be expected to remain within the ER via p40PX binding. Fusion to an ER-retention signal (STEVORss-p40PX-GFPSDEL) retains the protein in ER, co-localization with anti-ERC. Scale bar, 5 mm.(d) Localization of Hrs-GFP to the cytoplasm and membranes of food vacuole (labelled anti-CRT) and apicoplast (labelled anti-ACP). STEVORss-Hrs-GFP localizes to the ER as shown by co-localization with anti-ERC; however, mutation of residues necessary for lipid binding in both Hrs FYVE fingers (STEVORss-HrsC215S-GFP) does not abrogate ER localization, demonstrating PI(3)P-independent retention in ER. Addition of ER retention signal (STEVORss-Hrs-GFPSDEL) also causes ER localization. Scale bar, 5 mm. Boddey JA, O'Neill MT, Lopaticki S, Carvalho TG, Hodder AN, Nebl T, Wawra S,van West P, Ebrahimzadeh Z, Richard D, Flemming S, Spielmann T, Przyborski J,Babon JJ, Cowman AF. Export of malaria proteins requires co-translationalprocessing of the PEXEL motif independent of phosphatidylinositol-3-phosphatebinding. Nat Commun. 2016 Feb 1;7:10470.
See original on MMP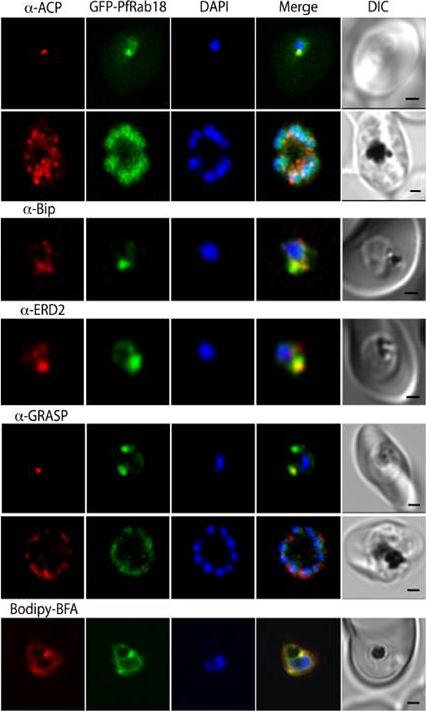
GFP-Rab18 fluorescence co-localizes with ER/Golgi, dense granules or microneme markers. GFP-PfRab1A fluorescence is distinct from the localization of markers for the apicoplast (ACP), the ER (Bip), the Golgi (ERD2 and GRASP), as well as from staining of the ER/Golgi with Bodipy BFA.Morse D, Webster W, Kalanon M, Langsley G, McFadden GI. Plasmodium falciparum Rab1A Localizes to Rhoptries in Schizonts. PLoS One. 2016 11(6):e0158174
See original on MMP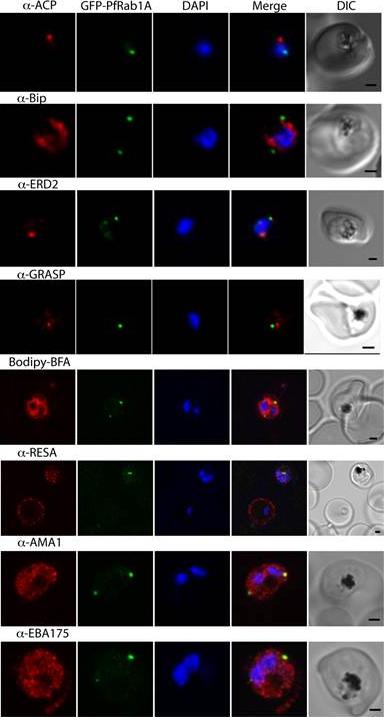
GFP-PfRab1A fluorescence does not co-localize with ER/Golgi, dense granules or microneme markers. GFP-PfRab1A fluorescence is distinct from the localization of markers for the apicoplast (ACP), the ER (Bip), the Golgi (ERD2 and GRASP), as well as from staining of the ER/Golgi with Bodipy BFA. GFP-Rab1A fluorescence is also distinct from the localization of markers for dense granules (RESA) or micronemes (AMA1 and EBA175).Morse D, Webster W, Kalanon M, Langsley G, McFadden GI. Plasmodium falciparum Rab1A Localizes to Rhoptries in Schizonts. PLoS One. 2016 Jun 27;11(6):e0158174
See original on MMP
Select gene expression constructs not targeted to the apicoplast are now co-localised with the endoplasmic reticulum. TMDs are represented by boxes and are joined by loops. All PfoTPT constructs are episomally expressed under the PfCRT promoter and tagged with triple HA at the C-terminus detected with anti-HA and secondary antibody conjugated to FITC (green). The parasite ER is detected with anti-BiP (red). Nuclei are stained with Hoechst (blue), and transmitted light images of the parasites within their host red blood cell are shown at right. Scale bars = 2μm. A. Point mutation of tyrosine residue at position 10 (✪) in the synthetic PfoTPT (Y10A) relocates protein from the apicoplast (B) to the ER, as well as the plasma. B. Removal of TMD 10 (TMD1to9), which completely abrogates targeting to the apicoplast (C), results in perinuclear localisation of the protein that almost entirely overlaps with the ER marker, BiP. C. Removal of TMDs 3, 4, 5, 6, 7, 8, 9 and 10 (TMD1to2), which completely abrogates targeting to the apicoplast (G), results in re-localisation of most of the protein to the ER. D. Removal of TMDs 2, 3, 4, 5, 6, 7, 8, 9 and 10 (TMD1), which completely abrogates targeting to the apicoplast (H), primarily results in localisation of most of the protein to the ER. E. Recombining TMD1 with TMD10 (TMD1+10) was unable to reconstitute apicoplast targeting (I) and resulted in some perinuclear ER targeting, plus some parasite plasma membrane targeting.Lim L, Sayers CP, Goodman CD, McFadden GI. Targeting of a Transporter to the Outer Apicoplast Membrane in the Human Malaria Parasite Plasmodium falciparum. PLoS One. 2016 11(7):e0159603.
See original on MMPMore information
| PlasmoDB | PKNH_0906200 |
| GeneDB | PKNH_0906200 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | PK11_0590c, PKH_090580 |
| Orthologs | PBANKA_0938400 , PCHAS_0905900 , PF3D7_1108600 , PVP01_0909400 , PVX_091105 , PY17X_0940900 |
| Google Scholar | Search for all mentions of this gene |