PF3D7_1407800 plasmepsin IV (PM4)
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. falciparum 3D7 |
Possible |
15491999 | Theo Sanderson, Wellcome Trust Sanger Institute | |
| P. falciparum 3D7 |
Possible |
USF piggyBac screen (Insert. mut.) | USF PiggyBac Screen | |
| P. berghei ANKA |
Possible |
RMgm-808 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-314 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-315 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-316 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
PlasmoGEM (Barseq) | PlasmoGEM | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. falciparum 3D7 | Asexual |
Attenuated |
15491999 | Theo Sanderson, Wellcome Trust Sanger Institute |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-808
Asexual blood stages show a (slightly) reduced growth rate and reduced hemozoin formation. |
Imported from RMgmDB |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-316
Asexual blood stages show a slightly reduced growth rate and show a virulence-attenuated phenotype in mice (see 'Additional remarks phenotype'). |
Imported from RMgmDB |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-314
Asexual blood stages show a slightly reduced growth rate and show a virulence-attenuated phenotype in mice (see 'Additional remarks phenotype'). |
Imported from RMgmDB |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-315
Asexual blood stages show a slightly reduced growth rate and show a virulence-attenuated phenotype in mice (see 'Additional remarks phenotype'). |
Imported from RMgmDB |
| P. berghei ANKA | Asexual |
Attenuated |
PlasmoGEM (Barseq) | PlasmoGEM |
| P. berghei ANKA | Gametocyte |
No difference |
RMgm-316 | Imported from RMgmDB |
| P. berghei ANKA | Ookinete |
No difference |
RMgm-316 | Imported from RMgmDB |
| P. berghei ANKA | Oocyst |
No difference |
RMgm-316 | Imported from RMgmDB |
| P. berghei ANKA | Sporozoite |
No difference |
RMgm-316 | Imported from RMgmDB |
| P. berghei ANKA | Liver |
No difference |
RMgm-316 | Imported from RMgmDB |
Imaging data (from Malaria Metabolic Pathways)
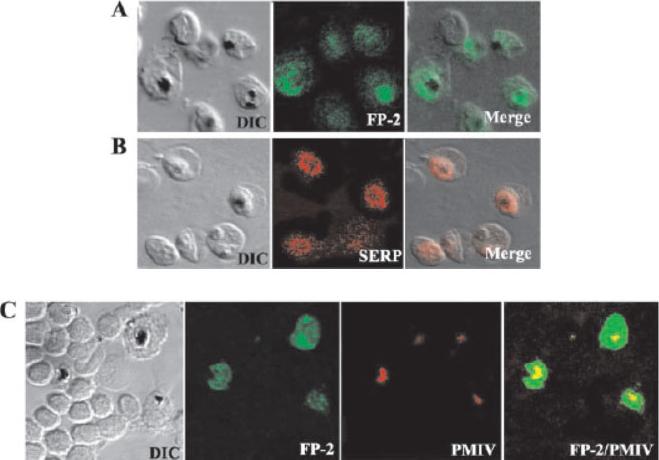
Immunolocalization of FP-2 by confocal microscopy. Erythrocytes infected with P. falciparum trophozoites were smeared on a glass slide, fixed with ice-cold methanol, and incubated with polyclonal FP-2 (A) or SERP (B) antibodies followed by Alexa 488 (green) or Alexa 594 (red) conjugated goat anti-rabbit IgG. Merged images of differential interference contrast (DIC) and fluorescence microscopy are shown on the right. C and D, slides were incubated with polyclonal FP-2 together with either plasmepsin IV (C) antibodies followed by Alexa-488-conjugated (green) goat anti-rabbit IgG and Alexa 594-conjugated (red) goat anti-mouse IgG. Merged images of green and red micrographs are shown on the right. The FP-2 antibody stained the entire trophozoite and vesicles or vesicle-like structures of unknown identity were seen extending into the erythrocyte cytoplasm. Co-staining of infected erythrocytes with antibodies to plasmepsin IV, which is present exclusively in the food, confirmed that FP-2 is present in the food vacuole as well as in the area that lies outside of the food vacuole (C). A comparison of FP-2 and SERP staining shows that the FP-2 is localized in the PVM as well as in a compartment that is distinct from the PVM.Dhawan S, Dua M, Chishti AH, Hanspal M. Ankyrin peptide blocks falcipain-2-mediated malaria parasite release from red blood cells. J Biol Chem. 2003 278:30180-6
See original on MMP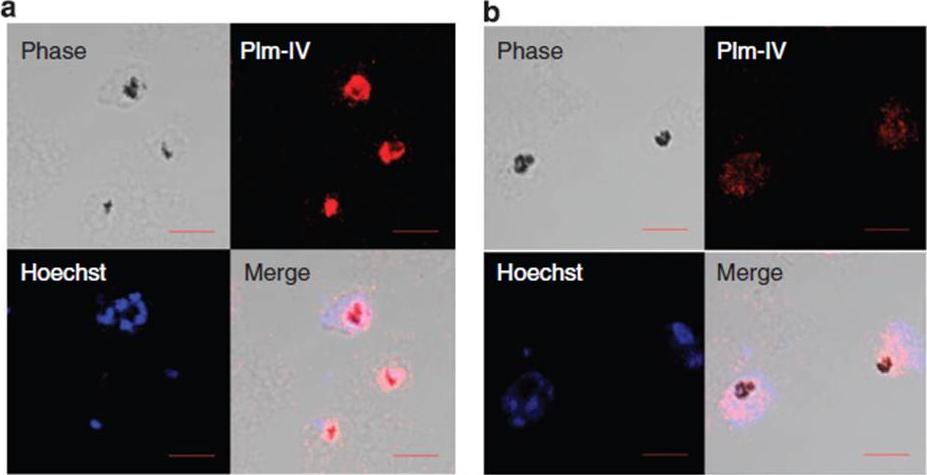
Plm-IV localization assay. (a) Confocal microscopy images show that Plm-IV co-localizes with the dark hemozoin-containing DV in healthy 3D7 P. falciparum parasites but not in (b) parasites treated with 3 mM of CQ for 4 h, where Plm-IV staining is less intense and is found throughout the parasite cytoplasm. (Scale bar set at 5 mm).Ch'ng JH, Lee YQ, Gun SY, Chia WN, Chang ZW, Wong LK, Batty KT, Russell B, Nosten F, Renia L, Tan KS. Validation of a chloroquine-induced cell death mechanism for clinical use against malaria. Cell Death Dis. 2014 Jun 26;5:e1305.
See original on MMPMore information
| PlasmoDB | PF3D7_1407800 |
| GeneDB | PF3D7_1407800 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | PF14_0075 |
| Orthologs | PBANKA_1034400 , PCHAS_1035200 , PKNH_1350300 , PVP01_1340900 , PVX_086040 , PY17X_1036800 |
| Google Scholar | Search for all mentions of this gene |