PF3D7_1335100 merozoite surface protein 7 (MSP7)
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. falciparum 3D7 |
Possible |
18820076 | Theo Sanderson, Wellcome Trust Sanger Institute | |
| P. berghei ANKA |
Possible |
RMgm-673 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-671 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-224 | Imported from RMgmDB | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. falciparum 3D7 | Asexual |
Attenuated |
18820076 | Theo Sanderson, Wellcome Trust Sanger Institute |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-673
msp7 infected BALB/c mice showed only a mild delay in the parasite growth rate during the initial phase of infection. The mice progressively developed a high parasitemia and died during the third week post infection similarly to wild type-infected mice.In C57BL/6 mice, msp7 parasites showed a significant reduction of the growth rate up to day 12 post infection and failed to induce signs and symptoms of ECM. These mice showed a prolonged course of infection with increasing parasitemia and died during the fourth week post infection with a peak parasitemia reaching 70%. All wild type-infected mice died at around day 8 post-infection when the parasitemia was still low (around 20%) developing a typical ECM pathology. The virulence attenuated phenotype of msp7 parasites varied with the mouse strain used. A substantial fraction of infected msp7 CD1 mice (about 60%) died within 10 days post-infection with signs and symptoms of ECM and with a parasitemia below 15% while the others mice died later of severe anemia. |
Imported from RMgmDB |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-671
Mutant blood stages show a minor growth defect resulting in a lower growth rate in BALB/c mice during the initial phase of infection (day 8/10). At later stages of infection (when reticulocytes are prevalent), the growth rate was similar to that of wild type parasites. In aged WISTAR rats (15 weeks) the mutant blood stages showed a reduced growth and virulence phenotype. |
Imported from RMgmDB |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-224
Mutants show a minor growth defect resulting in a lower growth rate in Balb/c mice during the initial phase of infection. When reticulocytes were prevalent (after day 9 after infection), the growth rate was similar to that of wild type parasites |
Imported from RMgmDB |
Imaging data (from Malaria Metabolic Pathways)
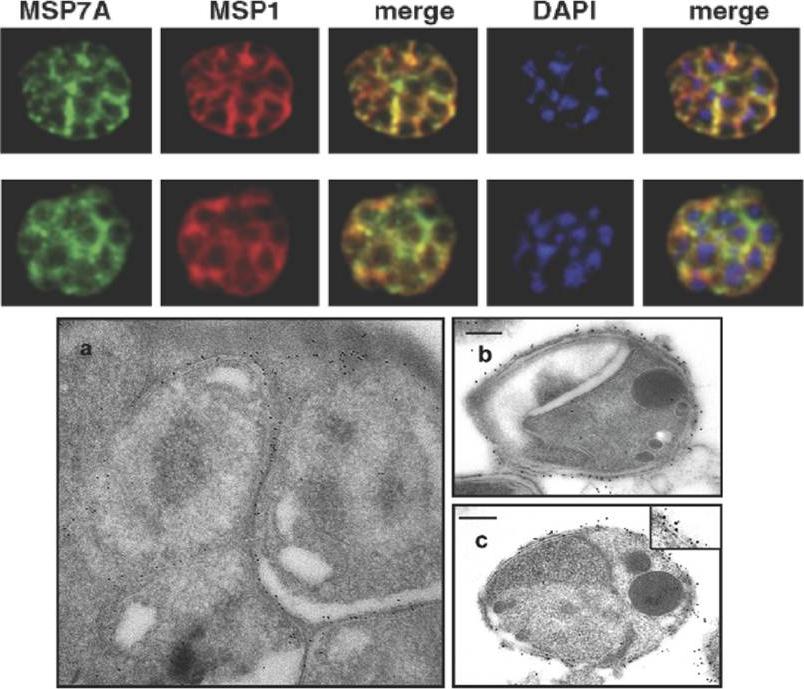
MSP7 co-localizes with MSP1 PFI1475w on mature blood stages and merozoites of P. falciparum. Acetone–methanol fixed mature P. falciparum T9/96 schizonts were analysed by IFA and visualised with mouse anti-MSP7A (MSP7A) and rabbit anti-MSP1 (MSP1) antibodies. DAPI was used to stain nuclei. A merge of anti-MSP7 and anti-MSP1, and of anti-MSP7 and anti-MSP1 and DAPI are shown. (D) Transmission electron micrograph showing MSP7-specific immuno-gold staining of the surface of (a) developing merozoites within the schizont and (b) a free merozoite; (c) double immuno-gold staining of the surface of a free merozoite labelled for MSP7 (the smaller, 5 nm gold particles) and MSP1 (the larger, 10 nm gold particles). The inset in (c) shows double labelling of part of the surface at higher magnification. Scale bar = 200 nm. Pachebat JA, Kadekoppala M, Grainger M, Dluzewski AR, Gunaratne RS, Scott-Finnigan TJ, Ogun SA, Ling IT, Bannister LH, Taylor HM, Mitchell GH, Holder AA. Extensive proteolytic processing of the malaria parasite merozoite surface protein 7 during biosynthesis and parasite release from erythrocytes. Mol Biochem Parasitol. 2007 51:59-69. Copyright Elsevier 2009.
See original on MMP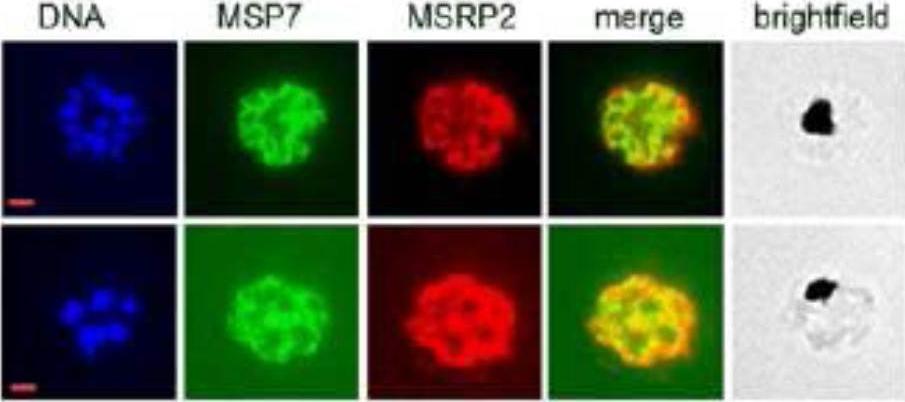
Co-localization of MSP7 and MSRP2. Late-stage parasites were formaldehyde-fixed, acetone-permeabilised and reacted with mAb 2G10, (mouse monoclonal anti-MSP7 antibody) and rabbit polyclonal anti-MSRP2 antibodies in an immunofluorescence assay.Kadekoppala M, Ogun SA, Howell S, Gunaratne RS, Holder AA. A systematic genetic analysis of the Plasmodium falciparum MSP7-like family reveals differences in protein expression, location and importance in asexual growth of the blood stage parasite. Eukaryot Cell. 2010 9:1064-74.
See original on MMP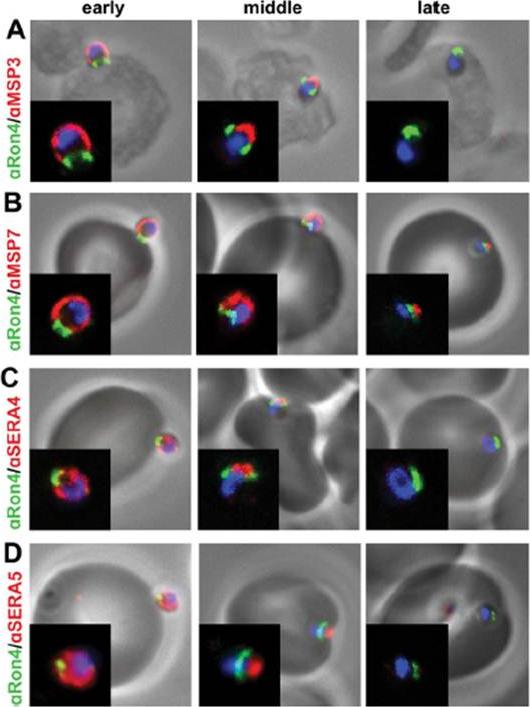
Peripherally associated proteins MSP3, MSP7, SERA4, and SERA5 are shed during invasion. Invading merozoites were labeled with PfRON4 as a marker of the tight junction (green) and colabeled with antibodies directed against MSP3 (A), MSP7 (B), SERA4 (C), or SERA5 (D). All proteins were shed at the point of tight junction, with antibody reactivity to the merozoite surface being observed only on the proportion of the merozoite external to the RBC. All primary antibodies were used at a 1:100 serum dilution, and secondary antibodies were used at a 1:500 dilution.Boyle MJ, Langer C, Chan JA, Hodder AN, Coppel RL, Anders RF, Beeson JG. Sequential processing of merozoite surface proteins during and after erythrocyte invasion by Plasmodium falciparum. Infect Immun. 2014 82(3):924-36.
See original on MMP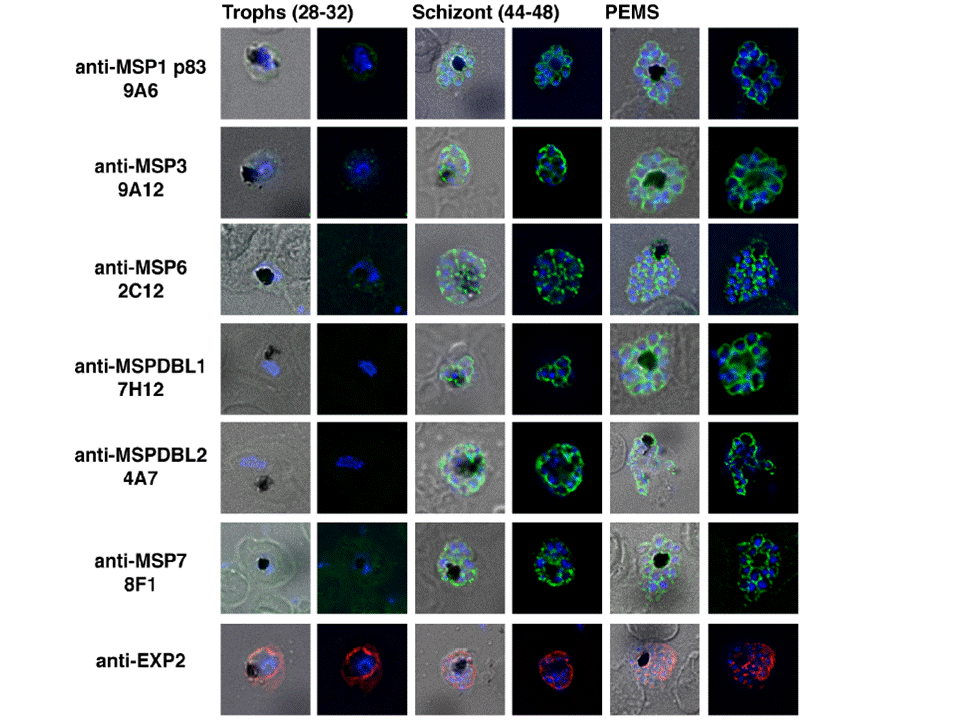
Processed Peripheral MSPs become part of an anchored complex late in schizogony. Immunofluorescence assays of Trophozoites (28-32 hr) in columns 1 (Phase + 488 + DAPI) and 2 (488 + DAPI), Schizonts (44-48 hr) in columns 3 (Phase + 488 + DAPI) and 4 (488 + DAPI) and PEMS (E64 treated) in columns 5 (Phase + 488 + DAPI) and 6 (488 + DAPI) were probed with monoclonal antibodies (green); 9A6 (anti-MSP1 p83), 9A12 (anti-MSP3), 2C12 (anti-MSP6), 7H12 (anti-MSPDBL1), 4A7 (anti-MSPDBL2) and 8F1 (anti-MSP7). EXP2 was used as an expression control probed with polyclonal rabbit anti-EXP2 antibodies (red). The nuclei of parasites are stained with DAPI (blue).Lin CS, Uboldi AD, Epp C, Bujard H, Tsuboi T, Czabotar PE, Cowman AF. Multiple P. falciparum Merozoite Surface Protein 1 Complexes Mediate Merozoite Binding to Human Erythrocytes. J Biol Chem. 2016 Jan 28. [Epub ahead of print] PMID:
See original on MMPMore information
| PlasmoDB | PF3D7_1335100 |
| GeneDB | PF3D7_1335100 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | PF13_0197 |
| Orthologs | PBANKA_1349100 , PCHAS_1353700 , PKNH_1266000 , PVP01_1219700 , PVP01_1220000 , PVP01_1220200 , PVX_082655 , PVX_082665 , PVX_082680 , PY17X_1354100 |
| Google Scholar | Search for all mentions of this gene |