PF3D7_1200600 erythrocyte membrane protein 1, PfEMP1 (VAR2CSA)
Disruptability [+]
| Species | Disruptability | Reference | Submitter |
|---|---|---|---|
| P. falciparum 3D7 |
Possible |
16025132 Does not bind CSA |
Theo Sanderson, Wellcome Trust Sanger Institute |
| P. falciparum 3D7 |
Possible |
USF piggyBac screen (Insert. mut.) | USF PiggyBac Screen |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. falciparum 3D7 | Asexual |
Altered cytoadherence |
16025132 Does not bind CSA |
Theo Sanderson, Wellcome Trust Sanger Institute |
Imaging data (from Malaria Metabolic Pathways)
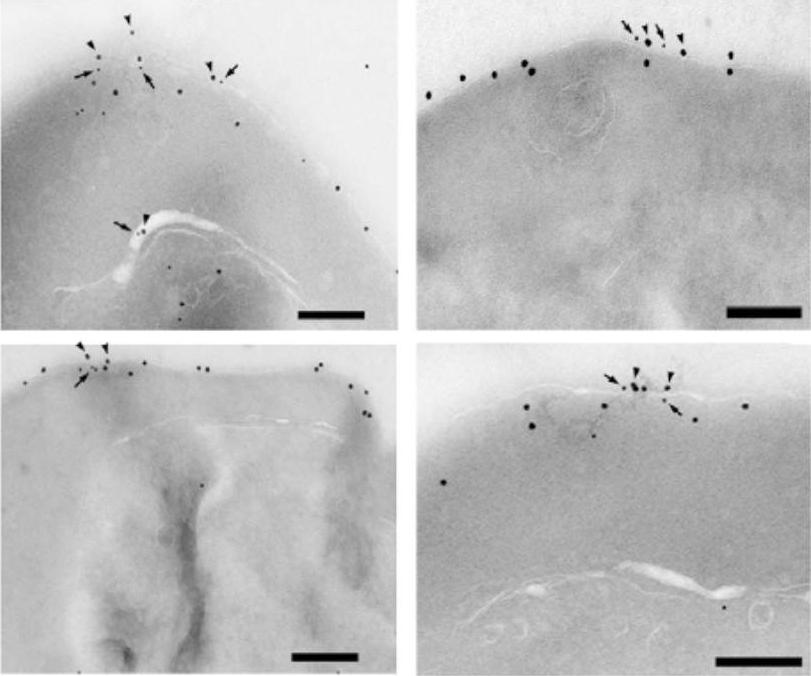
VAR2CSA and CLAG9 are present in the IRBC membranes of C4S-adherent parasites. The C4S-adherent FCR3 parasites, at the late ring to early trophozoite stages, were analyzed by immunoelectron microscopy by using rat anti-VAR2CSA and rabbit anti-CLAG9 primary antibodies followed by biotin-conjugated goat anti-rat IgG and 18-nm gold particle-conjugated goat anti-rabbit IgG secondary antibodies. The bound biotin-labeled antibodies were probed with 12-nm gold particles conjugated streptavidin. Arrows and arrowheads indicate, respectively, 12-and 18-nm gold particles. (Scale bars: 200 nm.) a substantial level of CLAG9 is located mainly at or in close proximity of the IRBC membrane in association with VAR2CSA.Goel S, Valiyaveettil M, Achur RN, Goyal A, Mattei D, Salanti A, Trenholme KR, Gardiner DL, Gowda DC. Dual stage synthesis and crucial role of cytoadherence-linked asexual gene 9 in the surface expression of malaria parasite var proteins. Proc Natl Acad Sci U S A. 2010 107:16643-8.
See original on MMP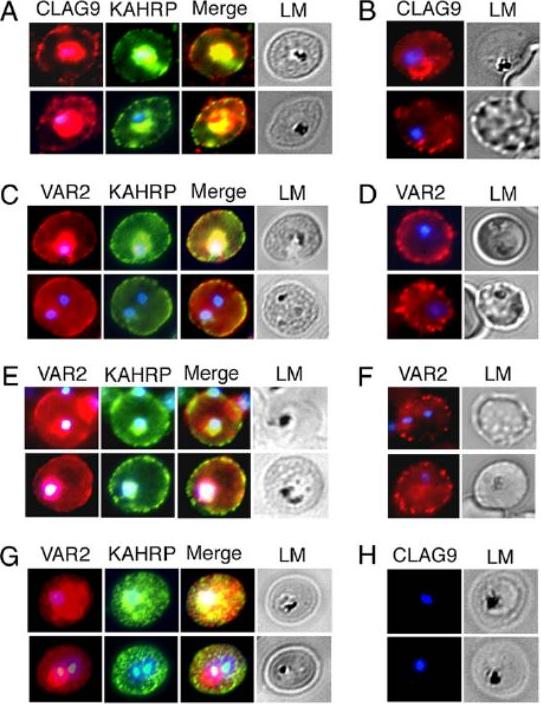
VAR2CSA and CLAG9 localize predominantly to IRBC membranes of C4S-adherent parasites. Immunofluorescence and light micrographs (LM) of membrane-permeabilized (A and C) and intact (B and D) IRBCs of C4S-adherent FCR3 parasites, harvested at the late ring/early trophozoite stage, were stained by using rabbit anti-CLAG9 (A and B), rabbit anti-VAR2CSA (C and D), and mouse anti-KAHRP (control) antibodies at dilutions given in SI Materials and Methods. (E and F) Immunofluorescence and LM of permeabilized (E) and intact (F) C4S-adherent 3D7 IRBCs stained as above with anti-VAR2CSA antibodies. (G and H), Immunofluorescence and LM of permeabilized clag9−/− parasites stained with anti-VAR2CSA antibodies (G) and anti-CLAG9 antibodies (H). Anti-CLAG9 antibodies showed a strong staining of the IRBC membrane (A). A substantial level of CLAG9 is located at or in close proximity to the erythrocyte membrane. Nonpermeabilized IRBCs also showed onsiderable levels of staining of the erythrocyte surface with anti-CLAG9 antibodies (B). Anti-VAR2CSA antibodies also localized VAR2CSA on the erythrocyte membrane of both permeabilized and nonpermeabilized IRBCs of the C4S-adherent parasites (C and D)Goel S, Valiyaveettil M, Achur RN, Goyal A, Mattei D, Salanti A, Trenholme KR, Gardiner DL, Gowda DC. Dual stage synthesis and crucial role of cytoadherence-linked asexual gene 9 in the surface expression of malaria parasite var proteins. Proc Natl Acad Sci U S A. 2010 107:16643-8.
See original on MMP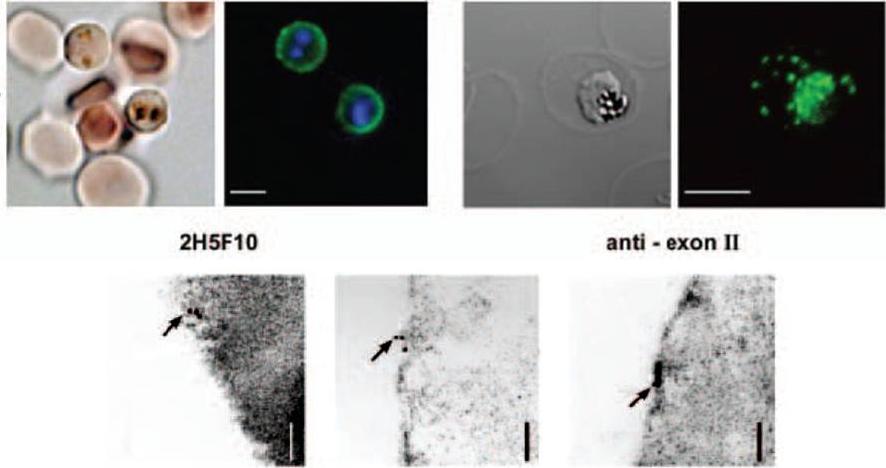
Characterization of the PfEMP1 variant located on the surface of erythrocytes infected with varCSA. Immunofluorescence of live infected erythrocytes using a monoclonal antibody recognizing the DBL3γ domain of PfEMP1CSA (2H5F10). An FITC-conjugated secondary antibody was used. In each case the left panel shows the light microscopy image, and the right panel an overlay of the fluorescent image and DNA staining using Hoechst 33324. Bar = 5 μm.Lower panel: Immunoelectron micrograph of fixed infected erythrocytes using an antisera recognizing the conserved PfEMP1 cytoplasmic domain. Gold particles are indicated by arrows. gold labelling in close association with the host erythrocyte plasma membrane. Bar = 120 nm.Andrews KT, Pirrit LA, Przyborski JM, Sanchez CP, Sterkers Y, Ricken S, Wickert H, Lépolard C, Avril M, Scherf A, Gysin J, Lanzer M. Recovery of adhesion to chondroitin-4-sulphate in Plasmodium falciparum varCSA disruption mutants by antigenically similar PfEMP1 variants. Mol Microbiol. 2003 49:655-69. Copyright John Wiley & Sons Ltd. 2010.
See original on MMP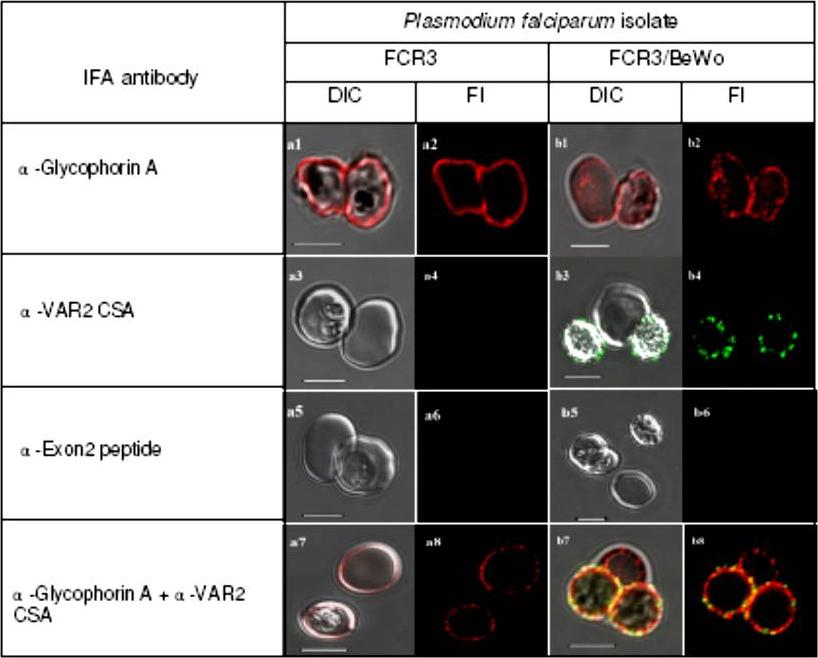
Specific fluorescent antibody labelling of surface antigens on live P. falciparum-infected erythrocytes selected for cytoadhesion. All plates in the odd numbered plates show the DIC shadow-cast image of with the fluorescence image superimposed. All even numbered plates show the fluorescence image alone. The a series shows antibody binding to unselected P. falciparum FCR3 infected erythrocytes. Plates in the b series show P. falciparum FCR3 infected erythrocytes which have been pre-selected for adhesion to BeWo cells. Before preparation for microscopy magnetically purified cells were seeded with control uninfected erythrocytes. The antibodies used in each row of images are indicated on the left. Scale bar 5 μM. Plates a1 and a2 show live staining of two intact infected erythrocytes of unselected FCR3 with the anti-Glycophorin A antibody, detected with Alexa®568-conjugated anti-mouse IgG. Glycophorin A staining appears uniform across the surface of these intact, infected erythrocytes. Plates a3 and a4 show the negative reaction of infected and uninfected erythrocytes with an antiserum specific for the VAR2CSA protein. Plates a5 and a6 show that intact, infected erythrocytes cannot be stained with an antibody recognizing the Exon 2-encoded intracellular domain of PfEMP1 proteins. Plates a7 and a8 show that incubating unselected FCR3 with both anti-glycophorin A and anti-VAR2CSA antibodies results in only glycophorin A fluorescence. The reaction is negative with the anti-VAR2CSA antiserum because this PfEMP1 antigen is not expressed by the great majority (99.5%) of unselected FCR3 cells. Specific detection of the VAR2CSA PfEMP1 on FCR3/BeWo cells is also shown. Both infected (plate b1, right cell), and uninfected erythrocytes (plate b1, left cell) are intact and recognized by the anti-glycophorin A antibody. Plates b3 and b4 illustrate the specific reactivity of the anti-VAR2CSA antibody with the surface of FCR3/BeWo cells. Plates b5 & b6 show that the intra-cellularPfEMP1 domain is, as expected, not accessible to the anti-Exon 2 antiserum in live, infected erythrocytes (the bottom right cell in plate b5 is uninfected). Plates b7 & b8 show that co-incubating FCR3/BeWo with anti-glycophorin A and anti-VAR2CSA antibodies results in double staining of the infected erythrocytes but glycophorin A staining alone on the uninfected erythrocyte (centre top in plates b7 & b8).Bengtsson D, Sowa KM, Salanti A, Jensen AT, Joergensen L, Turner L, Theander TG, Arnot DE. A method for visualizing surface-exposed and internal PfEMP1 adhesion antigens in Plasmodium falciparum infected erythrocytes. Malar J. 2008 Jun 5;7:101.
See original on MMP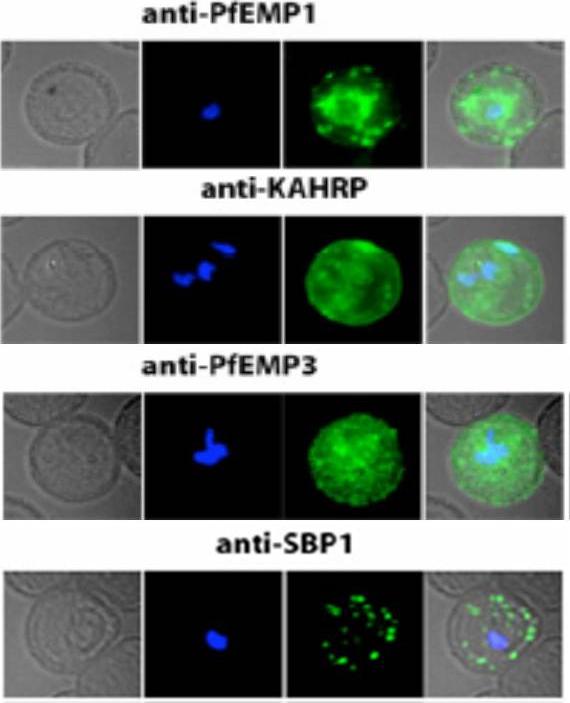
Localization of PfEMP1, KAHRP, PfEMP3 and SBP1 in P. falciparum-infected erythrocytes. The first column of each panel shows a bright-field image, the second panel specific antibody and the third panel overlay of the previous two and a nuclear stain (DAPI). PfEMP1 displays and functions on the surface of infected erythrocytes. KAHRP and PfEMP3 are exported to the erythrocyte cytoplasm and interacts with Maurer’s clefts and eventually reach the inner side of the host cell membrane. SBP1 is a Maurer’s clefts resident protein.Maier AG, Rug M, O'Neill MT, Brown M, Chakravorty S, Szestak T, Chesson J, Wu Y, Hughes K, Coppel RL, Newbold C, Beeson JG, Craig A, Crabb BS, Cowman AF. Exported proteins required for virulence and rigidity of Plasmodium falciparum-infected human erythrocytes. Cell. 2008 134:48-61.
See original on MMP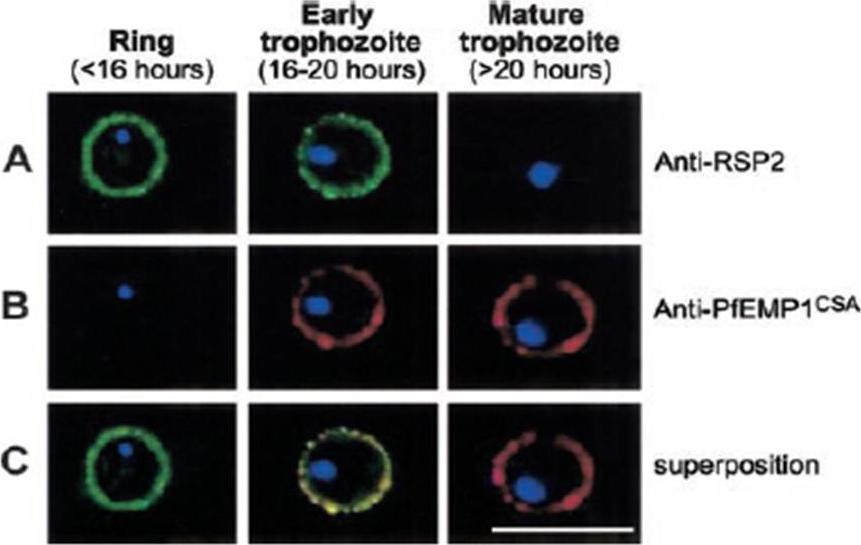
Expression profile of RSP2, the N terminal part of the Rap2 gene, and PfEMP1 at the erythrocyte surface during the blood-stage cycle. RSP-2CSA was stained with mAb B4 (green) and PfEMP1CSA with mAb 1B4/D4 (red) by L-IFA on IE. Parasite nuclei are stained by DAPI (blue). (A) Positive anti–RSP2 staining on synchronized ring stage–infected erythrocytes (16 hours after invasion) and early trophozoites (16 to 20 hours after invasion). No surface staining was detectable on mature forms (> 20 hours after invasion). (B) Absence of PfEMP1 staining on rings (16 hours) but strong IFA signal with early trophozoite and mature stages. (C) Superposition of anti–RSP2 and anti-PfEMP1 staining shows colocalization in early trophozoite stages. Scale bar measures 10 mmDouki JB, Sterkers Y, Lépolard C, Traoré B, Costa FT, Scherf A, Gysin J. Adhesion of normal and Plasmodium falciparum ring-infected erythrocytes to endothelial cells and the placenta involves the rhoptry-derived ring surface protein-2. Blood. 2003 101:5025-32.
See original on MMP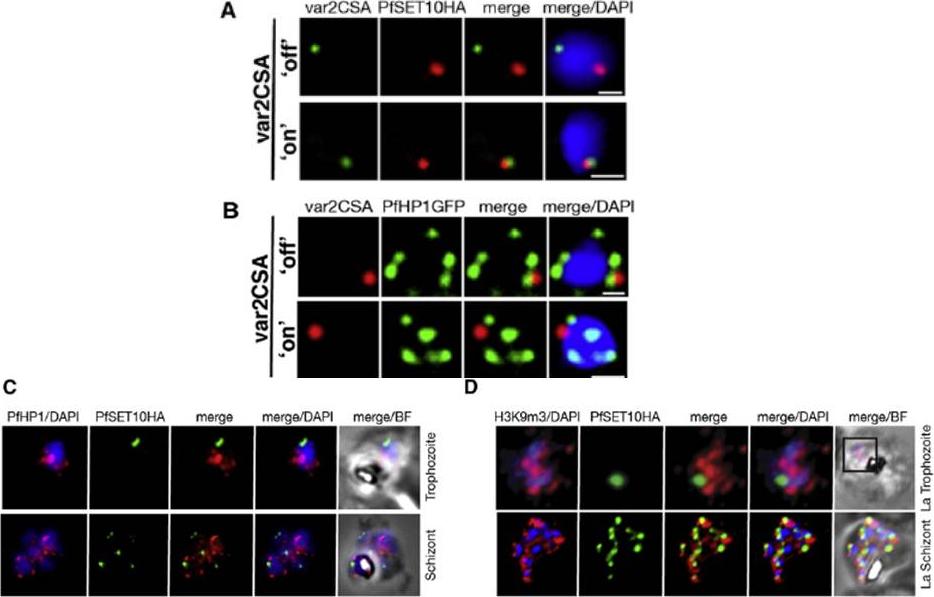
A. IFA-FISH of 3D7SET10-HA, in which var2csa (PFL0030c) activation has been selected by CSA adherence (‘‘on’’) (bottom panel). Unselected 3D7SET10-HA (top panel) has var2csa ‘‘off.’’ The localization of var2csa (green) and PfSET10 (red) is shown with both merged with the nucleus stained by DAPI (blue). Scale bars are equivalent to 0.5 mm. B. PfHP1 does not localize with active var2csa (‘‘on’’). The var2csa gene (red) location is shown with respect to PfHP1 protein (green). C. PfHP1 (red) and D. histone mark H3K9me3 (red,), known markers of silent var gene clusters and PfSET10 (green), do not colocalize in the nucleus stained with DAPI (blue).Volz JC, Bártfai R, Petter M, Langer C, Josling GA, Tsuboi T, Schwach F, Baum J, Rayner JC, Stunnenberg HG, Duffy MF, Cowman AF. PfSET10, a Plasmodium falciparum Methyltransferase, Maintains the Active var Gene in a Poised State during Parasite Division. Cell Host Microbe. 2012 11(1):7-18.
See original on MMP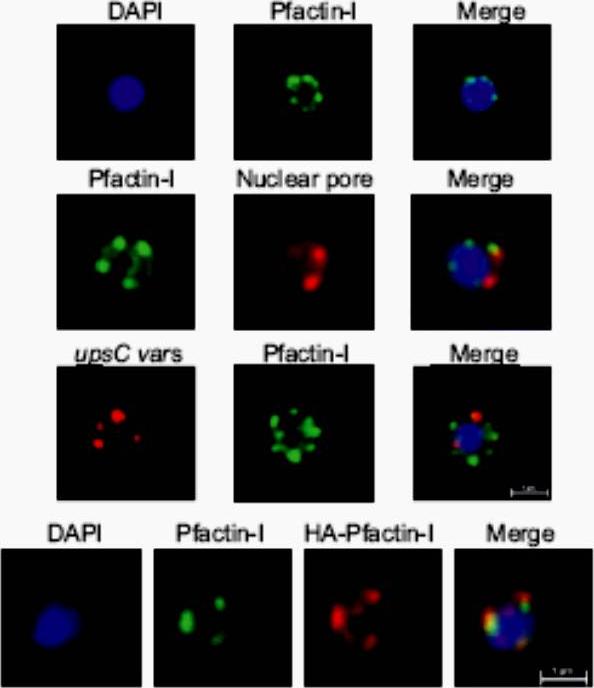
Upper panel: IFA visualization of subcellular actin in rings using an anti-Pfactin-I antibody (rabbit #2). Second panel: Two-color IFA assay using anti-Pfactin-I (rabbit #2, green) and anti-nuclear pore (PF14_0706) antibody (rat, red). Third panel: Combined FISH/IFA assay in rings showing partial colocalization of chromosome internal var genes (upsC-type) and actin. Fourth panel: IFA analysis of HA-actin-transfected ring stage parasites using mouse anti-HA antibody (red) and anti-Pfactin-I antibody (rabbit #2, green). IFA analysis with anti-Pfactin-I antibodies or with anti-HA antibody in HAactin-transfected 3D7 parasites confirmed the restricted location of actin to the nucleus in ring stage parasites and demonstrated that it concentrated in foci at the nuclear periphery. Colocalization with a nuclear pore antibody indicates that the staining is limited to the inner side of the nucleus.Zhang Q, Huang Y, Zhang Y, Fang X, Claes A, Duchateau M, Namane A, Lopez-Rubio JJ, Pan W, Scherf A. A critical role of perinuclear ilamentous actin in spatial repositioning and mutually exclusive expression of virulence genes in malaria parasites. Cell Host Microbe. 2011 10:451-63.
See original on MMP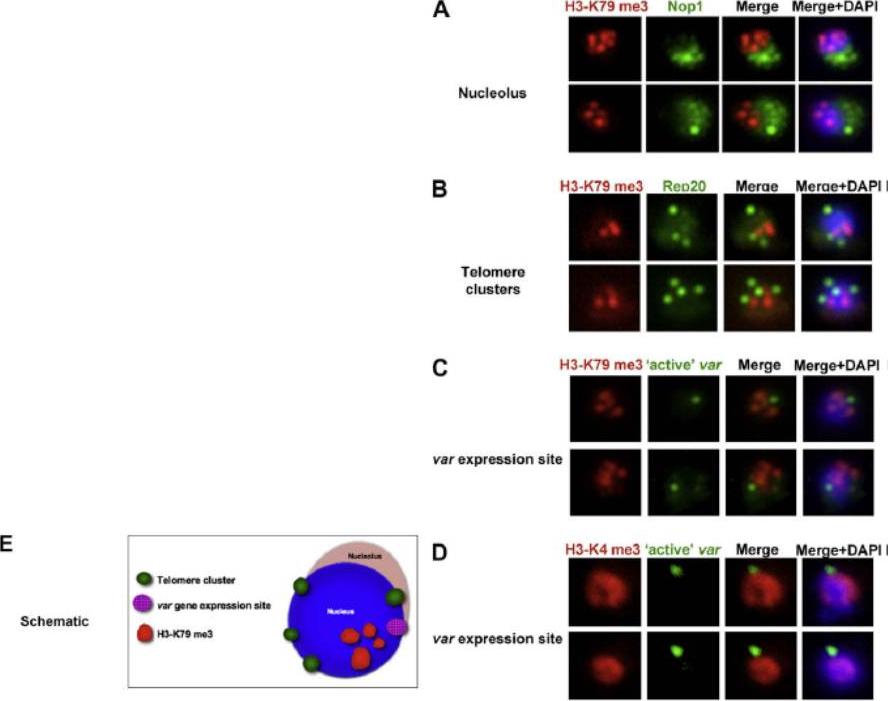
Co-localisation of H3-K79me3 with known cellular compartments. A. Nucleolus: Double labelling IFAs with antibodies against H3-K79me3 and nucleolus marker, PfNop1. B. Telomere clusters: IFeFISH analysis between H3-K79me3 (red) and Rep20 (green). C. Active var gene: IF-FISH analysis between H3-K79me3 (red) and active var gene (green). D. Active var gene colocalisation with activating mark, H3-K4me3. IF-FISH analysis between H3-K4me3 (red) and active var gene (green). DAPI stained nucleus in blue. Font colour represents corresponding fluorescein labelled antibody. Scale bar depicts 1 mm. E. Schematic of the dynamic compartmen-talisation of the P. falciparum nucleus. The distinct sub-nuclear distribution of histone methylation modification, taking example of H3-K79me3 as observed in this study. Known compartments compatible with active transcription are indicated. H3-K79me3 occupies a distinct zone.Issar N, Ralph SA, Mancio-Silva L, Keeling C, Scherf A. Differential sub-nuclear localisation of repressive and activating histone methyl modifications in P. falciparum. Microbes Infect. 2009 11:403-7.
See original on MMP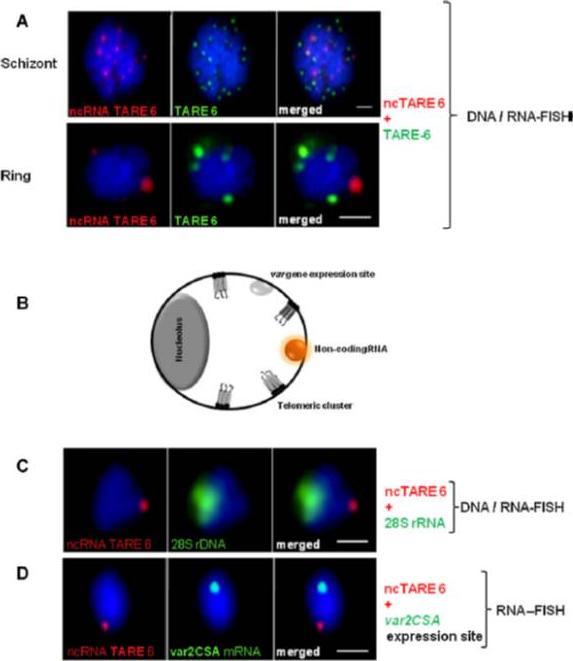
The ncRNAs define a novel subdomain in the P. falciparum nucleus. (A) RNA-FISH signals are shown in red, and DNA-FISH signals are shown in green (TARE-6). The signals of the TARE-6 ncRNA and telomeric DNA do not co-localize in ring stages, but in the late-schizont stage some telomere clusters are transcribed and co-localize with TARE-6 ncRNA probe. (B) P. falciparum nucleus present a complex nuclear architecture with several subcompartments localized in the nuclear periphery as are: telomeric clusters, var gene expression site, nucleolus and non-coding RNA compartment. (C) RNA-FISH signals are shown in red (TARE-6), and 28S rDNA DNA-FISH signals are in green. The signals from 28S rDNA and the non-coding TARE-6 transcript do not co-localize. (D) Two-color RNA-FISH of a var2CSA RNA probe (green) and TARE-6 ss ncRNA transcripts (red) for ring stage parasites (FCR3-CSA parasite population). The signals from the var2CSA transcripts and the TARE-6 ncRNAs do not co-localize. In all images, nuclear DNA was stained with DAPI (blue). Scale bars: 1 mm for the ring and schizont stages.Sierra-Miranda M, Delgadillo DM, Mancio-Silva L, Vargas M, Villegas-Sepulveda N, Martínez-Calvillo S, Scherf A, Hernandez-Rivas R. Two long non-coding RNAs generated from subtelomeric regions accumulate in a novel perinuclear compartment in Plasmodium falciparum. Mol Biochem Parasitol. 2012 185(1):36-47. Copyright Elsevier.
See original on MMP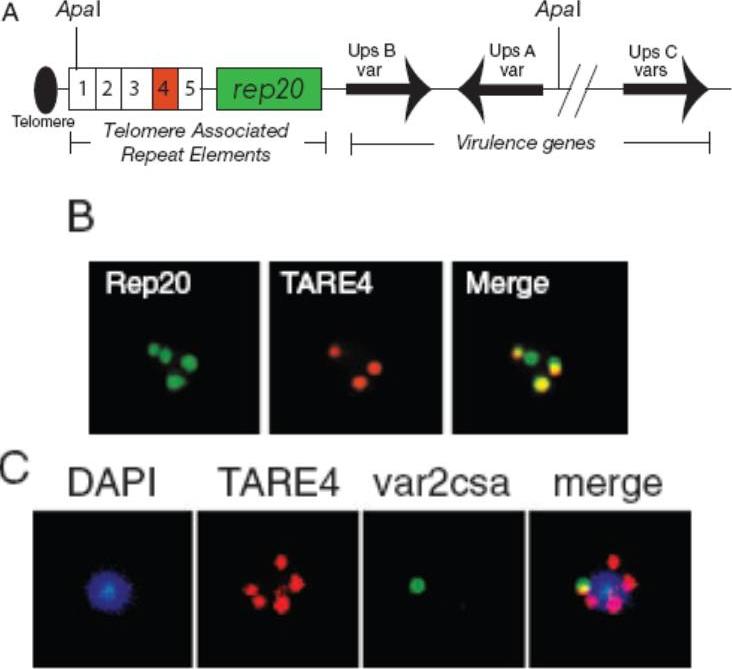
A. Schematic of a P. falciparum chromosome end. Chromosome ends are comprised of the parasite’s virulence genes, the telomere and six non-coding TAREs. This structure is conserved across the subtelomeric regions. FISH probes were made to Rep20 (TARE6, shaded green), the largest and most conserved of these repeats, and TARE4 (shaded red), a much smaller, highly degenerate repeat. ApaI sites used in the 2D-PFGE analysis are indicated. B. FISH telomeric probes are specific. Epifluorescence image of a nucleus hybridized with a Rep20 (green) and TARE4 (red) telomeric probes. The telomeric probes colocalize. The var gene var2csa colocalizes with telomeric clusters irrespective of its transcriptional status. C. FISH analysis on intact nuclei. Epifluorescence images of a nucleus stained with DAPI and hybridized with var2csa (green) and TARE4 (red) probes to identify var2csa and telomeric clusters respectively.Marty AJ, Thompson JK, Duffy MF, Voss TS, Cowman AF, Crabb BS. Evidence that Plasmodium falciparum chromosome end clusters are cross-linked by protein and are the sites of both virulence gene silencing and activation. Mol Microbiol. 2006 62:72-83. Copyright John Wiley & Sons Ltd. 2010
See original on MMP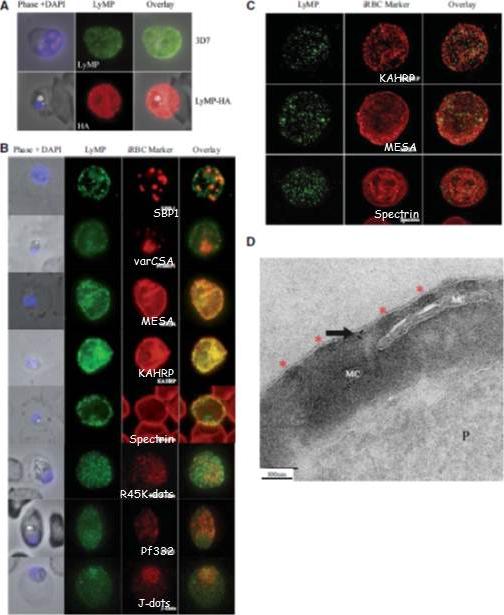
Localization of LyMP in iRBCs. A) Localization of LyMP in trophozoites, with either anti-LyMP (green) or anti-HA (red) showing that LyMP is exported into the RBC cytosol. 4’,6’-Diamidino-2-phenylidole (DAPI) is a nuclear stain (blue). B) Colocalization of LyMP (green) with a variety of iRBC markers (red) that are indicated in the panel. DAPI is a nuclear stain (blue). C) High-resolution structured illumination fluorescence microscopy (OMX) with anti-LyMP (green) and either anti-KAHRP, anti-MESA, or anti-spectrin (red) colocalizations. Corresponding overlay images are shown for all. D) Immunoelectron micrograph of gold-labeled Ha-tagged parasites. Red asterisks indicate knobs and the arrow indicates gold labeling of LyMP at the RBC membrane skeleton. P, parasite; MC, Maurer’s clefts. LyMP showed a pattern of distribution similar to that for MESA, KAHRP. LyMP did not consistently colocalize with MCs or to other punctate structures previously identified in the iRBC cytosol such as those seen for FIKK4.2 (R45) or for J-dots (KAHsp40).Proellocks NI, Herrmann S, Buckingham DW, Hanssen E, Hodges EK, Elsworth B, Morahan BJ, Coppel RL, Cooke BM. A lysine-rich membrane-associated PHISTb protein involved in alteration of the cytoadhesive properties of Plasmodium falciparum-infected red blood cells. FASEB J. 2014 Apr 4. [Epub ahead of print]
See original on MMP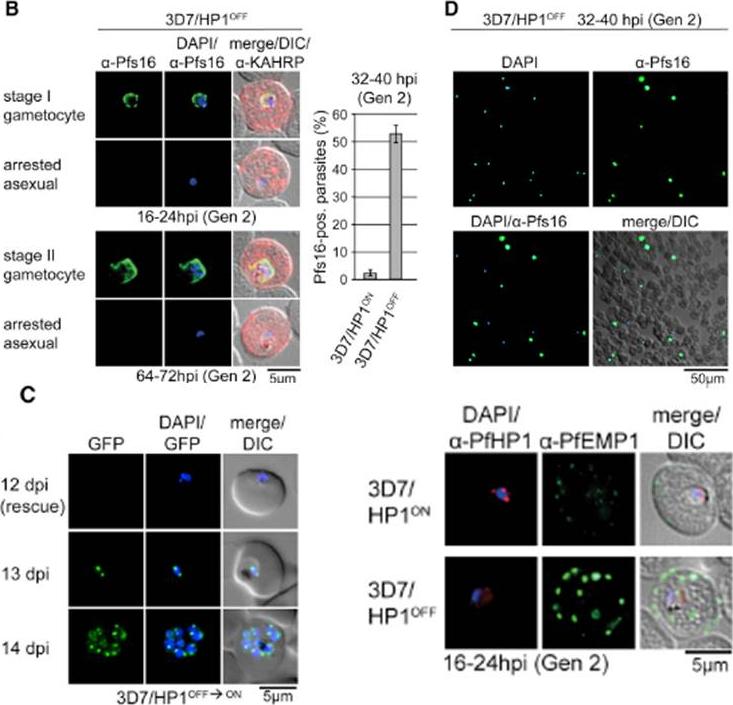
PfHP1 depletion induces gametocyte conversion. Distinction between 3D7/HP1OFF early gametocytes and arrested trophozoites by IFA (left) and proportion of Pfs16/KAHRP-positive parasites in 3D7/HP1ON and 3D7/HP1OFF (right). Values show the mean ± SD of three biological replicates (100 KAHRP-pos. iRBCs were scored per experiment). (D) a-Pfs16 IFA of a 3D7/HP1OFF parasite culture (Shield-1 removal at 4–12 hpi) at 32–40 hr postreinvasion (image taken at 40X magnification). The gametocyte hyperinduction phenotype is highlighted by the high proportion of Pfs16-positive stage I gametocytes among all DAPI-positive iRBCs. (C) Growth-arrested 3D7/HP1OFF parasites reenter mitotic proliferation after Shield-1 replenishment. dpi, days postreinvasion. PfHP1 depletion causes reversible cell-cycle arrest at the G1/S transition phase. Lower right panel: a-PfHP1/a-PfEMP1 (mAb 6H1) IFAs of 3D7/HP1ON and 3D7/HP1OFF parasites at 16–24 hpi in generation 2. PfEMP1 in 3D7/HP1OFF parasites at the single-cell level is hyperexpressed and correctly trafficked to the iRBC surfaceBrancucci NM, Bertschi NL, Zhu L, Niederwieser I, Chin WH, Wampfler R, Freymond C, Rottmann M, Felger I, Bozdech Z, Voss TS. Heterochromatin protein 1 secures survival and transmission of malaria parasites. Cell Host Microbe. 2014 16(2):165-76.
See original on MMP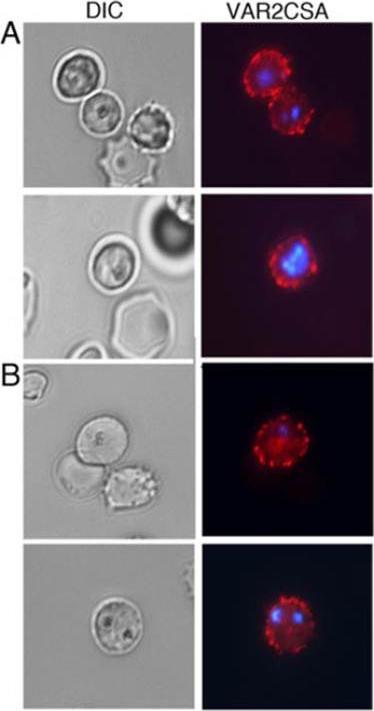
PFB0080c-knockout P. falciparum express VAR2CSA on the surface of infected erythrocytes. A and B, Immuno-fluorescence analysis of VAR2CSA in live ΔPFB0080c IRBCs was performed using rabbit anti-VAR2CSA antiserum followed by biotin-conjugated sheep antirabbit IgG secondary antibody and phycoerythrin-conjugated streptavidin. Shown are the immunofluorescent and light micrographs of parental 3D7-CSA parasites (A) and ΔPFB0080c parasites (B); both parasite types expressed VAR2CSA protein. Goel S, Muthusamy A, Miao J, Cui L, Salanti A, Winzeler EA, Gowda DC. Targeted disruption of a ring-infected erythrocyte surface antigen (RESA)-like export protein gene in Plasmodium falciparum confers stable chondroitin 4-sulfate cytodherence capacity. J Biol Chem. 2014 Oct 23. [Epub ahead of print]
See original on MMP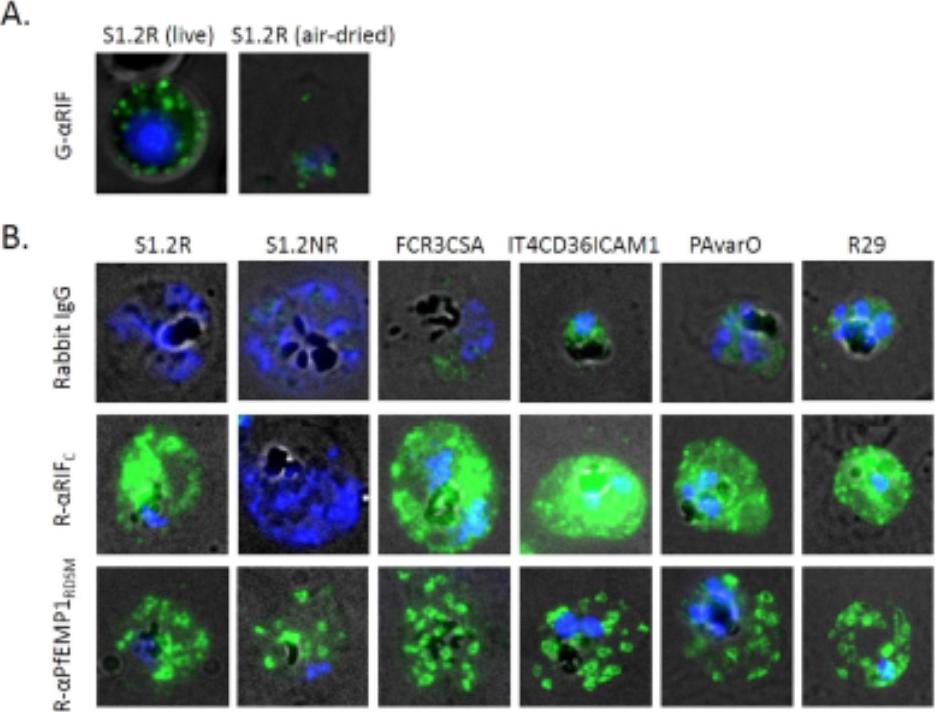
Indirect immunofluorescence micrographs. (A) Gα RIF IgG labelling of RIFINs on a live S1.2Rinfected erythrocyte and an air-dried iRBC. Unlike the antibody labeling of live S1.2R-infected cells which showed punctate surface-specific staining (left panel), air dried monolayers stained with Gα RIF IgG showed only a faint parasite cytoplasmic staining (right panel) suggesting that the native conformational (3D) epitope may have been lost during the fixing by desiccation. (B) Air-dried monolayers of erythrocytes infected with S1.2R, S1.2NR, FCR3CSA, IT4CD36ICAM1, PAvarO and R29 parasites stained with non-immune rabbit IgG (10 μ g/ml), anti-RIFIN Rα RIFC rabbit IgG (10 μg/ml) and Maurer’s cleft labelling by R-α PfEMP1RDSM IgG (8 μ g/ml). Alexa488-conjugated secondary anti-rabbit or anti-goat antibodies were used (1:200) as well as Vectashield with DAPI (nuclear staining). Merged channels of transmission light (black and white), green (Alexa 488) and blue (DAPI) and representative cells are shown here. Since desiccation results in breaks in the membrane and exposes the inner surface of the RBC membrane where the conserved RIFIN C-terminus is located, Rα RIFC IgG could readily label S1.2R-infected RBCs in a patchy manner. In comparison, IgG from Rα RIFI, R5α RIFC and R6α RIFC showed no notable staining (data not shown). Rα RIFC IgG was then applied to air-dried monolayers of other parasite strains to detect and localize potential A-RIFINs. While S1.2NR (Fig. 2B), NF54CSA and 3D7CD36ICAM1 (data not shown) had no notable staining, FCR3CSA, IT4CD36ICAM1, PAvarO and R29 parasites showed staining similar to S1.2 R.Ch'ng JH, Sirel M, Zandian A, Del Pilar Quintana M, Chun Leung Chan S, Moll K, Tellgren-Roth A, Nilsson I, Nilsson P, Qundos U, Wahlgren M. Epitopes of anti-RIFIN antibodies and characterization of rif-expressing Plasmodium falciparum parasites by RNA sequencing. Sci Rep. 2017 7:43190.
See original on MMPMore information
| PlasmoDB | PF3D7_1200600 |
| GeneDB | PF3D7_1200600 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | 2277.t00006, MAL12P1.6, PFL0030c |
| Orthologs | |
| Google Scholar | Search for all mentions of this gene |