PF3D7_1133400 apical membrane antigen 1 (AMA1)
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. falciparum 3D7 |
Refractory |
24571085 | Theo Sanderson, Wellcome Trust Sanger Institute | |
| P. falciparum 3D7 |
Possible |
USF piggyBac screen (Insert. mut.) | USF PiggyBac Screen | |
| P. berghei ANKA |
Possible |
RMgm-953 | Imported from RMgmDB | |
| P. berghei ANKA |
Refractory |
RMgm-10 | Imported from RMgmDB | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. falciparum 3D7 | Asexual |
Attenuated |
24571085 | Theo Sanderson, Wellcome Trust Sanger Institute |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-953
The AMA1ko mutant lacks expression of AMA1 and expresses GFP under the control of the constitutive hsp70 promoter. The mutant has not been cloned. Parasite populations have been analysed that consisted of mixtures of the AMA1ko GFP+ mutant parasites and parasites expressing AMA1 (AMA1+ GFP- parasites).Evidence is presented that the multiplication rate of AMA1ko GFP+ parasites is 35% that of wild type parasites. The AMA1ko GFP+ schizonts produced normal numbers of meorozoites, indicating that the decreased multiplication rate of AMA1KO parasites reflects a defect in merozoite entry into erythrocytes. |
Imported from RMgmDB |
| P. berghei ANKA | Sporozoite |
Difference from wild-type |
RMgm-953
The AMA1ko mutant lacks expression of AMA1 and expresses GFP under the control of the constitutive hsp70 promoter. The mutant has not been cloned. Parasite populations have been analysed that consisted of mixtures of the AMA1ko GFP+ mutant parasites and parasites expressing AMA1 (AMA1+ GFP- parasites).To test AMA1KO sporozoite capacity to invade hepatocytes, populations of AMA1ko GFP+/ AMA1+ GFP- parasites were transferred to mosquitoes. The same ratio of GFP+ versus GFP- sporozoites is found in the blood fed to mosquitoes and in the mosquito salivary glands, indicating that AMA1 has no detectable effect on parasite development in the mosquitoes. The capacity of these salivary gland sporozoites to invade cultured hepatocytes was then tested. After sporozoite incubation with HepG2 cells in vitro, a similar proportion of AMA1ko GFP+ versus AMA1+ GFP- parasites is found in the input sporozoites and in hepatic schizonts developing inside HepG2 cells 60 h post infection indicating that AMA1 has no role in P. berghei sporozoite infection of hepatocytes. |
Imported from RMgmDB |
| P. berghei ANKA | Liver |
Difference from wild-type |
RMgm-953
The AMA1ko mutant lacks expression of AMA1 and expresses GFP under the control of the constitutive hsp70 promoter. The mutant has not been cloned. Parasite populations have been analysed that consisted of mixtures of the AMA1ko GFP+ mutant parasites and parasites expressing AMA1 (AMA1+ GFP- parasites).To test AMA1KO sporozoite capacity to invade hepatocytes, populations of AMA1ko GFP+/ AMA1+ GFP- parasites were transferred to mosquitoes. The same ratio of GFP+ versus GFP- sporozoites is found in the blood fed to mosquitoes and in the mosquito salivary glands, indicating that AMA1 has no detectable effect on parasite development in the mosquitoes. The capacity of these salivary gland sporozoites to invade cultured hepatocytes was then tested. After sporozoite incubation with HepG2 cells in vitro, a similar proportion of AMA1ko GFP+ versus AMA1+ GFP- parasites is found in the input sporozoites and in hepatic schizonts developing inside HepG2 cells 60 h post infection indicating that AMA1 has no role in P. berghei sporozoite infection of hepatocytes. |
Imported from RMgmDB |
Imaging data (from Malaria Metabolic Pathways)
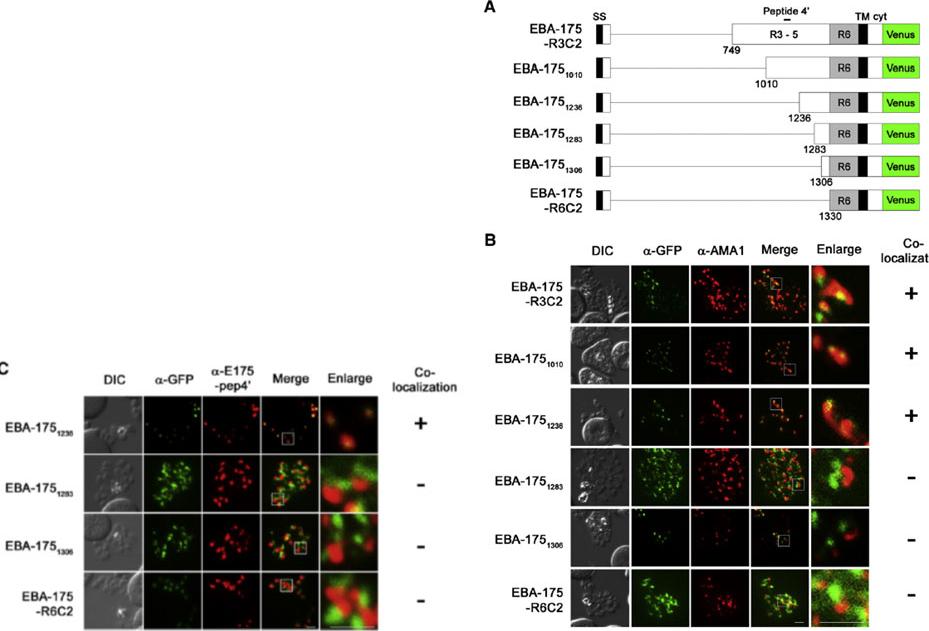
IFA detection of parasites expressing serially truncated recombinant EBA-175. (A) Schematic structures of recombinant EBA-175 proteins; EBA-175-R3C2, EBA-1751010, EBA-1751236, EBA-1751283, EBA-1751306 or EBA-175-R6C2. Signal sequence (SS), regions 1 to 6 (R1, R2, R3–5 and R6), transmembrane region (TM) and cytoplasmic tail (cyt) are indicated. Bar above the scheme indicates the target region of anti-EBA-175-pep4′ antibody (aa 1089–1108). (B and C) Parasites were stained with anti-GFP (α-GFP, green) and anti-AMA1 (panel B; α-AMA1, red) or anti-EBA-175-pep4′ (panel C; α-E175-pep4′, red). Scale bar indicates 2 μm. DIC is a differential interference contrast image. Status of the co-localization with a microneme marker is shown at the right side of each row. (+) or (−) indicate that two signal co-localized or not co-localized.Sakura T, Yahata K, Kaneko O. The upstream sequence segment of the C-terminal cysteine-rich domain is required for microneme trafficking of Plasmodium falciparum erythrocyte binding antigen 175. Parasitol Int. 2012 62(2):157-64
See original on MMP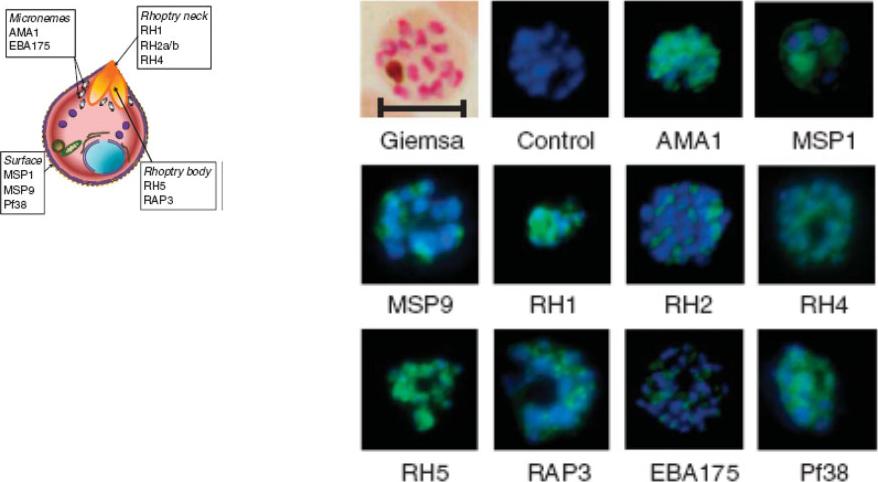
Indirect immunofluorescence images of P. falciparum schizonts stained with rabbit IgG (green) induced by 10 viral-vectored vaccines expressing malaria antigens, and negative control vectors lacking a malaria antigen. Nuclei are counterstained with 4,6-diamidino-2-phenylindole (blue). All sera were tested against 3D7 strain parasites, with the exception of anti-PfRH1 for which FVO strain parasites were used. All images to same scale as Giemsa stained image (top left, on which scale bar indicates 5 μm).Douglas AD, Williams AR, Illingworth JJ, Kamuyu G, Biswas S, Goodman AL, Wyllie DH, Crosnier C, Miura K, Wright GJ, Long CA, Osier FH, Marsh K, Turner AV, Hill AV, Draper SJ. The blood-stage malaria antigen PfRH5 is susceptible to vaccine-inducible cross-strain neutralizing antibody. Nat Commun. 2011 2:601.
See original on MMP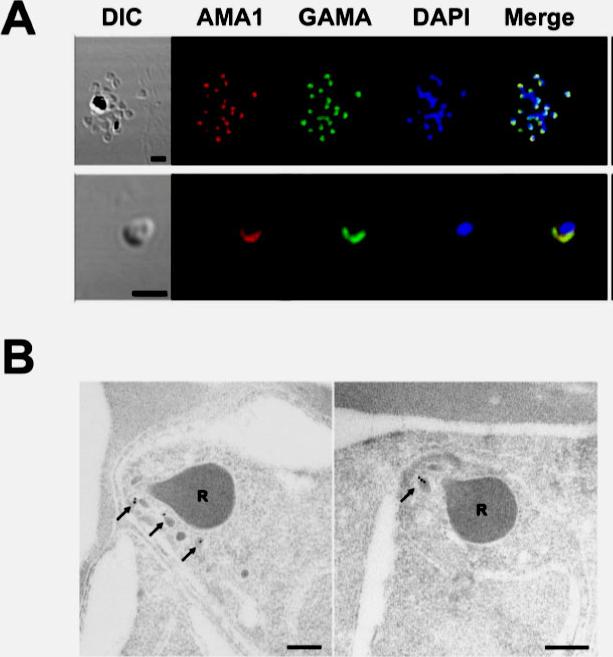
Localization of GAMA in asexual blood stage parasites. (A) GAMA localization using immuno-fluorescence assay. Acetone-fixed P. falciparum 3D7 mature schizonts (top panel) and free merozoites (bottom panel) were probed with rabbit anti-FL (green) and mouse anti-PfAMA1 (microneme marker)(red). Parasite nuclei were stained with DAPI (blue). Scale bars represent 2 μm. (B) GAMA localization using immunoelectron microscopy. The two sections of merozoites in schizont-infected erythrocytes were probed with purified rabbit anti-Tr3 antibody and subsequently by secondary antibody conjugated with gold particles. The arrows indicate the micronemal localization of signals from gold particles. Bars represent 200 nm. Arrows mark micronemes.Arumugam TU, Takeo S, Yamasaki T, Thonkukiatkul A, Miura K, Otsuki H, Zhou H, Long CA, Sattabongkot J, Thompson J, Wilson DW, Beeson JG, Healer J, Crabb BS, Cowman AF, Torii M, Tsuboi T. Discovery of GAMA, a Plasmodium falciparum merozoite micronemal protein, as a novel blood-stage vaccine candidate antigen. Infect Immun. 2011 79(11):4523-32
See original on MMP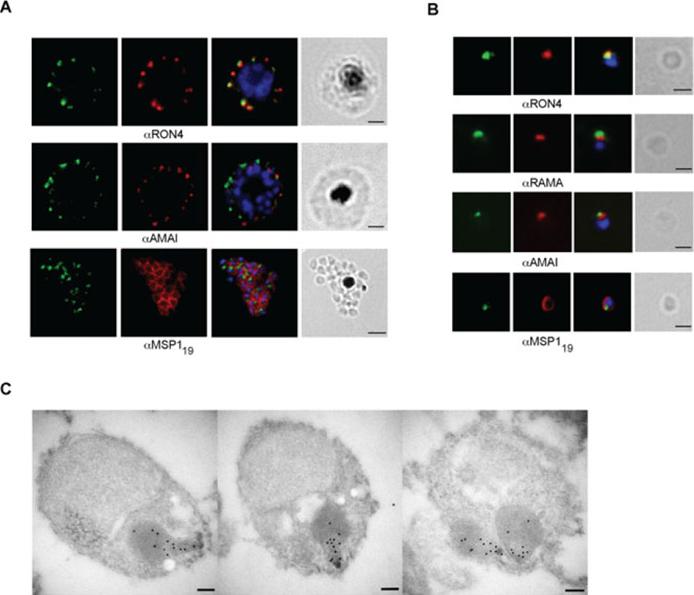
RON12 is located within the rhoptry neck.. A and B. Immunofluorescence and bright-field images of schizonts (A) and merozoites (B). Images from left to right show anti-RON12 labelling (green) followed by microneme (anti-AMA1), rhoptry (neck: anti-RON4; body: anti-RAMA) and surface (anti-MSP119) specific antibodies in red, overlay of both with DAPI-stained nuclei and the bright-field image. Size bars in (A) equal 2 μm and in (B) 1 μm. C. Immuno-electron microscopy of isolated merozoites. Localization of RON12 in the rhoptry neck of three different merozoites is shown. Size bars equal 100 nm.Knuepfer E, Suleyman O, Dluzewski AR, Straschil U, O'Keeffe AH, Ogun SA, Green JL, Grainger M, Tewari R, Holder AA. RON12, a novel Plasmodium-specific rhoptry neck protein important for parasite proliferation. Cell Microbiol. 2013 Aug 12. [Epub ahead of print]
See original on MMP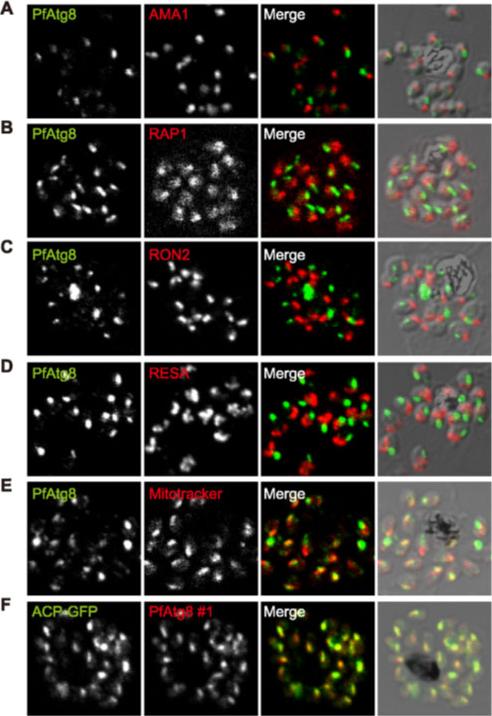
PfAtg8 localizes to the apicoplast. P. falciparum FCR3 (A–E) and P. falciparum 3D7 transfected with ACP-GFP (F–H) were stained with the indicated organelle markers and visualized by confocal microscopy (because ACP-GFP was not uniformly expressed, some merozoites displayed only faint GFP signals). Anti-PfAtg8 antibody #1 was used in (A–F), and anti-PfAtg8 antibody #2 was used in (G). Apical membrane antigen 1 (AMA1) as a microneme marker (A), rhoptry-associated protein 1 (RAP1) as a rhoptry body marker (B), rhoptry neck protein 2 (RON2) as a rhoptry neck marker (C), the ring-infected erythrocyte surface antigen (RESA) as a dense granule marker (D), MitoTrackerRed CMXRos as a mitochondria marker (E), ACPGFP (F–H) and the organellar histone-like protein PfHU (H) as an apicoplast marker were used. Scale bar, 1 mm.Kitamura K, Kishi-Itakura C, Tsuboi T, Sato S, Kita K, Ohta N, Mizushima N. Autophagy-Related Atg8 Localizes to the Apicoplast of the Human Malaria Parasite Plasmodium falciparum. PLoS One. 2012;7(8):e42977.
See original on MMP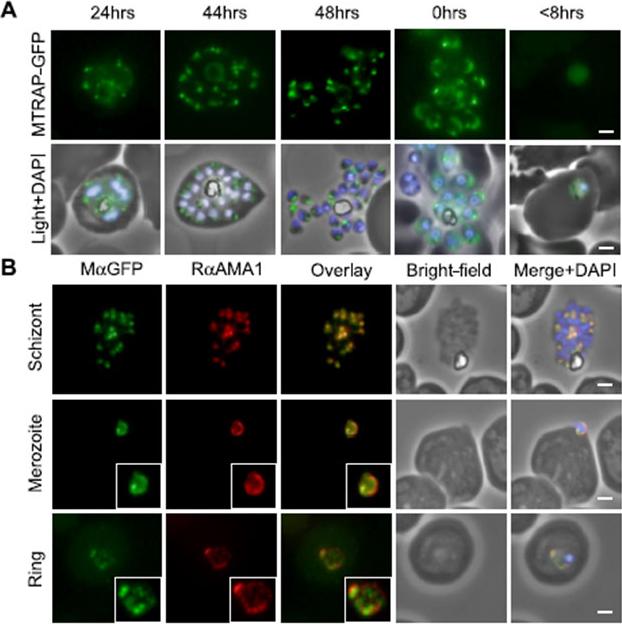
MTRAP undergoes extensive processing during invasion. A, episomally expressed MTRAP-GFP followed through the life cycle confirms apical localization (44–48 h) suggests the release of MTRAP onto the surface of free merozoites (capping, 0 h) and suggests that, despite processing, the cytoplasmic tail is carried through to newly invaded rings (<8 h). B, this is confirmed by following co-localization with antibodies that recognize AMA1, which have been previously reported as being carried through to rings. In early ring stages, there does not appear to be direct co-localization, suggesting that they may be associating with different compartments. Baum J, Richard D, Healer J, Rug M, Krnajski Z, Gilberger TW, Green JL, Holder AA, Cowman AF. A conserved molecular motor drives cell invasion and gliding motility across malaria life cycle stages and other apicomplexan parasites. J Biol Chem. 2006 281(8):5197-208.
See original on MMP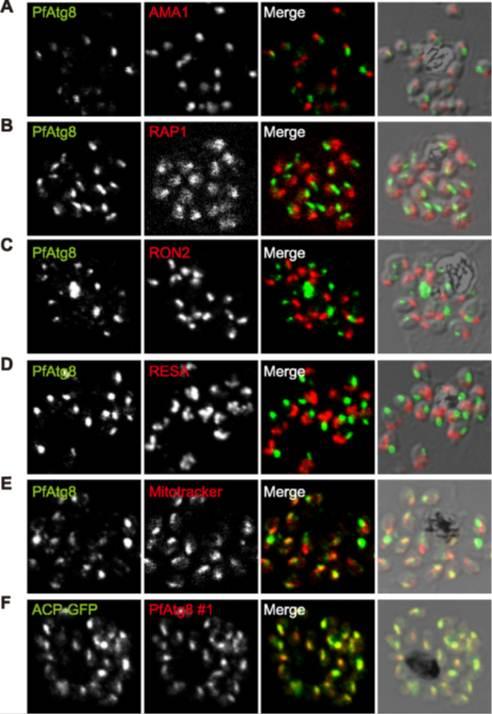
PfAtg8 localizes to the apicoplast. P. falciparum FCR3 (A–E) and P. falciparum 3D7 transfected with ACP-GFP (F–H) were stained with the indicated organelle markers and visualized by confocal microscopy (because ACP-GFP was not uniformly expressed, some merozoites displayed only faint GFP signals). Anti-PfAtg8 antibody #1 was used in (A–F), and anti-PfAtg8 antibody #2 was used in (G). Apical membrane antigen 1 (AMA1) as a microneme marker (A), rhoptry-associated protein 1 (RAP1) as a rhoptry body marker (B), rhoptry neck protein 2 (RON2) as a rhoptry neck marker (C), the ring-infected erythrocyte surface antigen (RESA) as a dense granule marker (D), MitoTrackerRed CMXRos as a mitochondria marker (E), ACPGFP (F–H) and the organellar histone-like protein PfHU (H) as an apicoplast marker were used. Scale bar, 1 mm.Kitamura K, Kishi-Itakura C, Tsuboi T, Sato S, Kita K, Ohta N, Mizushima N. Autophagy-Related Atg8 Localizes to the Apicoplast of the Human Malaria Parasite Plasmodium falciparum. PLoS One. 2012;7(8):e42977
See original on MMP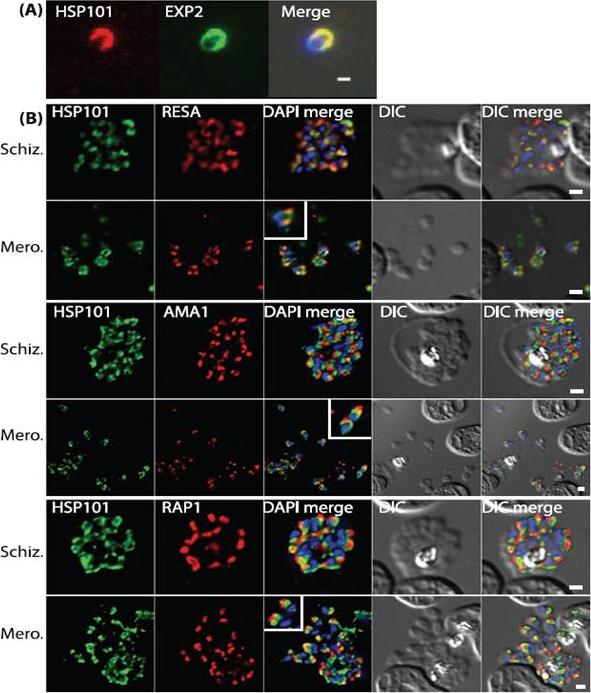
PTEX components are apically localised in merozoite dense granules. (A and B) Immunofluorescence assays were completed during schizogony and were probed for either PTEX components or markers of distinct organelles as indicated in each of the panels. (A) Merozoites probed with antibodies specific to both HSP101 and EXP2 demonstrate apical localization for both proteins. (B) Schizonts and merozoites were labeled with antibodies specific to HSP101 and either RESA (dense granules), AMA1 (micronemes) or RAP1 (rhoptries). Co-localisation quantitation reveals that HSP101 localises most closely with dense granule associated RESA (top two panels). Scale bar: 1μm. Bullen HE, Charnaud SC, Kalanon M, Riglar DT, Dekiwadia C, Kangwanrangsan N, Torii M, Tsuboi T, Baum J, Ralph SA, Cowman AF, de Koning-Ward TF, Crabb BS, Gilson PR. Biosynthesis, localisation and macromolecular arrangement of the Plasmodium falciparum translocon of exported proteins; PTEX. J Biol Chem. 2012 287(11):7871-84.
See original on MMP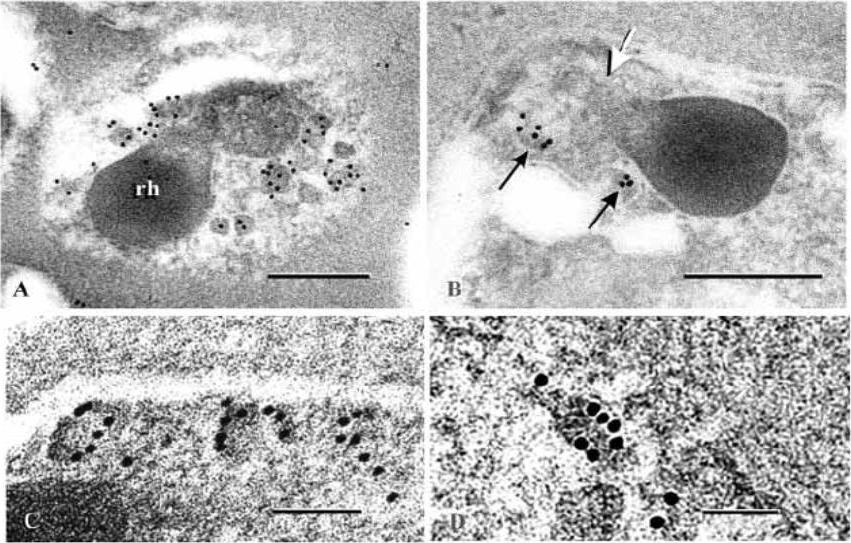
Immunogold labeling of maturing schizonts with antibody against PfAMA-1 whole ectodomain. (A) An obliquely sectioned merozoite apex with a cluster of micronemes strongly labelled by Pab 1, surrounding the tips of two unlabelled rhoptries (rh). Scale bar, 100 nm. (B) Two micronemes (black arrows) labelled by Pab 3 and a longitudinally sectioned rhoptry devoid of labeling, including its narrow apical end (white arrow). Scale bar, 100 nm. (C,D) PfAMA-1 labelling by Pab 3 around the peripheries of micronemes. The microneme in (D) was situated near the Golgi cisterna and represents an early stage in this organelle’s migration to the apex. Scale bars, 50 nm.Three polyclonal antibodies raised against the recombinant PfAMA-1 ectodomain sequence recognizing both the 83 kDa and processed 66 kDa molecules label the peripheries of translocating and mature micronemes but do not label rhoptries significantly at any stage of merozoite development within schizonts. This result confirms that PfAMA-1 is a micronemal protein, and indicates that within the microneme it is located near or inserted into this organelle’s boundary membrane.Adapted with permission from Bannister LH, Hopkins JM, Dluzewski AR, Margos G, Williams IT, Blackman MJ, Kocken CH, Thomas AW, Mitchell GH. Plasmodium falciparum apical membrane antigen 1 (PfAMA-1) is translocated within micronemes along subpellicular microtubules during merozoite development. J Cell Sci. 2003 116:3825-34. Copyright Elsevier
See original on MMP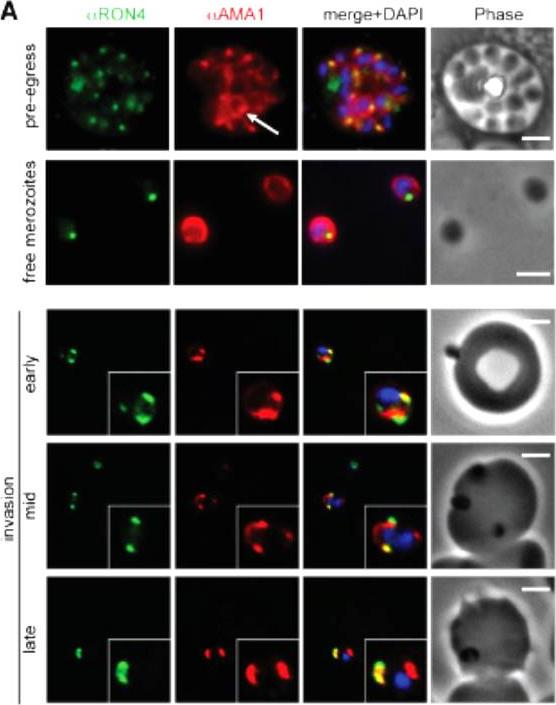
Wide field IFA time course of invasion from pre-egress through to invasion using anti-PfAMA1/PfRON4. Pre-egress and free merozoites show standard wide-field image. Invasion images show IFA with deconvolution (single slice). Scale bar = 2.0μm. In parasites fixed prior to or immediately after egress from E64-treated schizonts (ETS) a significant proportion of PfAMA1 had already been secreted onto the merozoite surface. Conventional fluorescence imaging of midinvasion merozoites revealed that PfAMA1 labeling localized directly within the plane of the ring of PfRON4 fluorescence. A proportion of PfAMA1 was also variably localized to the merozoite surface and unreleased micronemes.Riglar DT, Richard D, Wilson DW, Boyle MJ, Dekiwadia C, Turnbull L, Angrisano F, Marapana DS, Rogers KL, Whitchurch CB, Beeson JG, Cowman AF, Ralph SA, Baum J. Super-resolution dissection of coordinated events during malaria parasite invasion of the human erythrocyte. Cell Host Microbe. 2011 9:9-20.
See original on MMP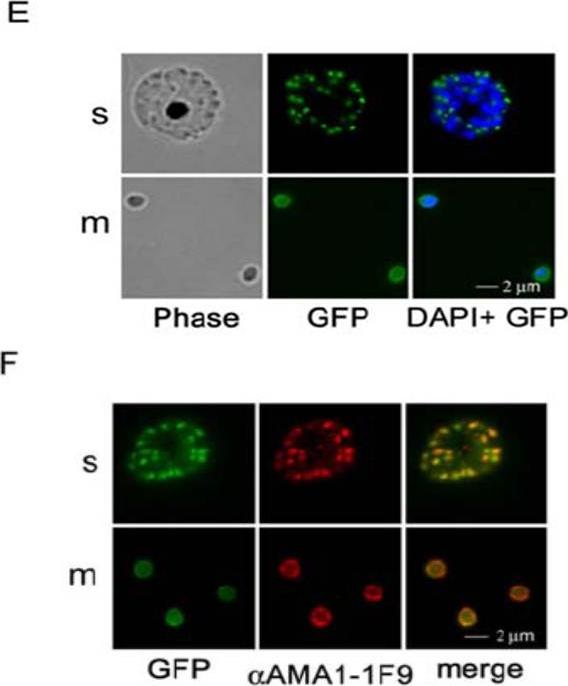
Expression and localization of 3D7- or W2mef-derived AMA-1 in transgenic parasites. Full-length AMA-1-GFP (green) localizes in schizonts (s) at the apical end of merozoites (m) and is distributed over the surface of merozoites after schizont rupture. This is shown in both unfixed (E) and fixed parasites (F). The distribution of the endogenous protein visualized with the 3D7-specific 1F9 antibody (red) is identical to the localization of the fusion protein (merge).Treeck M, Zacherl S, Herrmann S, Cabrera A, Kono M, Struck NS, Engelberg K, Haase S, Frischknecht F, Miura K, Spielmann T, Gilberger TW. Functional analysis of the leading malaria vaccine candidate AMA-1 reveals an essential role for the cytoplasmic domain in the invasion process. PLoS Pathog. 2009 Mar;5(3):e1000322.
See original on MMP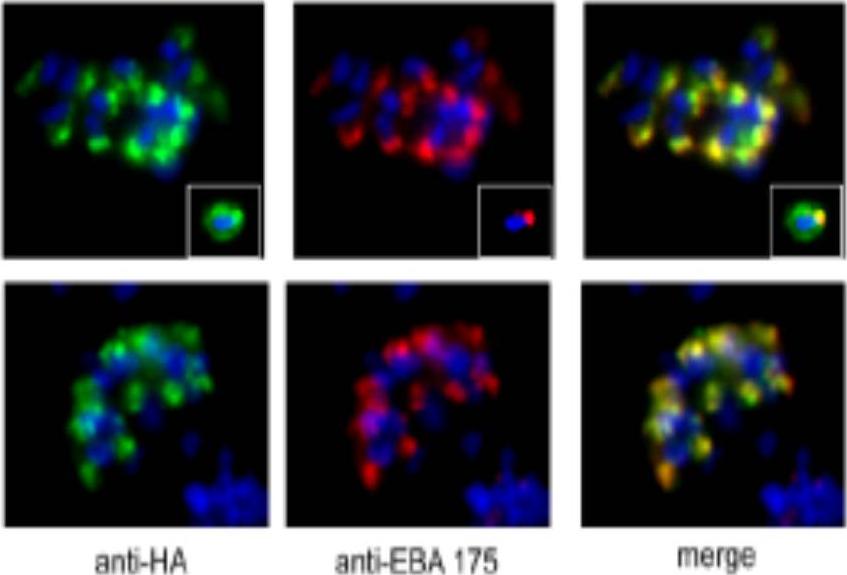
A. IFA of drug-selected 3D7 parasites harbouring the AMA_M3 construct. The transgenic PfAMA1/DIII-HA product detected by anti-HA mAb 3F10 (green) co-localises with the microneme protein EBA-175 (red) in mature segmented schizonts. The inset shows similar staining of a free merozoite, where some translocation of PfAMA1/DIIIHA onto the merozoite surface is detectable as is normal for endogenous PfAMA1. B. IFA of drug-selected 3D7 parasites harbouring the AMA_ins construct. The transgene product detected by anti-HA mAb 3F10 (green) co-localises correctly with the microneme protein EBA-175 (red) in mature segmented schizonts.Olivieri A, Collins CR, Hackett F, Withers-Martinez C, Marshall J, Flynn HR, Skehel JM, Blackman MJ. Juxtamembrane shedding of Plasmodium falciparum AMA1 is sequence independent and essential, and helps evade invasion-inhibitory antibodies. PLoS Pathog. 2011 7(12):e1002448.
See original on MMP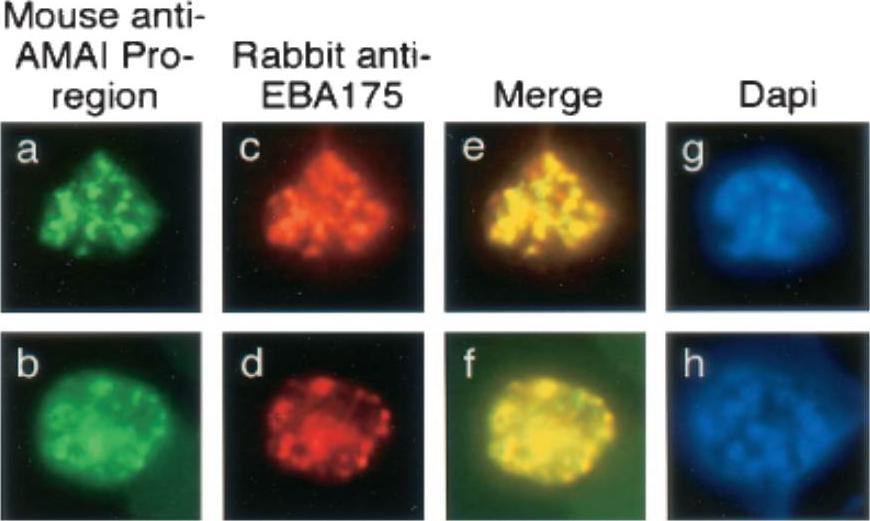
Single and dual indirect immunofluorescence images showing the same subcellular localization of AMA1 and EBA-175 in two mature schizonts of P. falciparum. (a to d) FITC-labeled MAb 5G8 against the PfAMA1 pro-region (a and b) and rhodamine-labeled anti-EBA175 (c and d). (e and f) Merged images show full-length AMA1 colocalized with EBA175. (g and h) DAPI-stained parasites included to show the stage of merozoite development. MAb 5G8, which specifically recognizes the unprocessed form of AMA1, showed a punctate pattern characteristic of the apical organelles of merozoites (a and b). Anti-EBA175 antibodies showed the same pattern (c and d), and merging of the images from the FITC and rhodamine channels suggested that these two proteins were colocalized in the same subcellular compartment, the micronemes.Healer J, Crawford S, Ralph S, McFadden G, Cowman AF. Independent translocation of two micronemal proteins in developing Plasmodium falciparum merozoites. Infect Immun. 2002 70:5751-8.
See original on MMP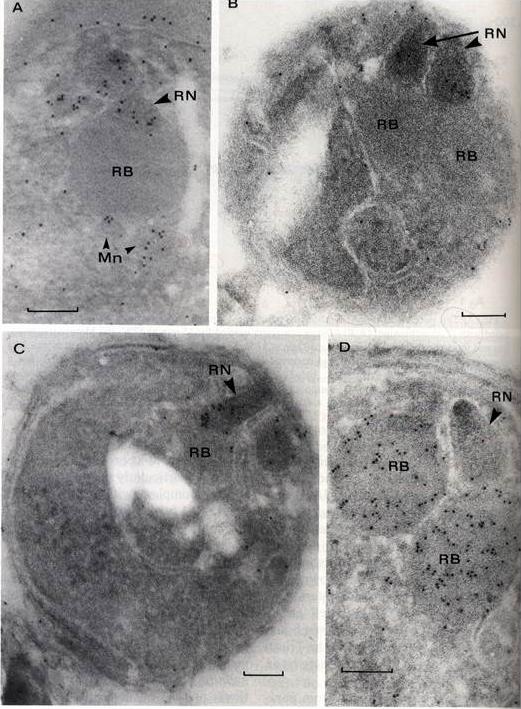
Post-embedding immunolableing of P. falciparum isolate FC27, with affinity-purified anti Ag352 antibodies to AMA-1 (A,B,C) and rabbit anti QF3 monoclonal antibody 7H8/50 to RAP-1 (D). Bars equal 100 nm. A. Maturing merozoites within a schizont showing gold particles over rhoptry neck (RN) and micronemes (Mn). B,C Free merozoites showing labeling of electron dense rhoptry necks (RN), no labeling over rhoptry bodies (RB), and sparse labeling of merozoite periphery. D. Immunolabeling restricted to the body of rhoptries (RB).Crewther PE, Culvenor JG, Silva A, Cooper JA, Anders RF. Plasmodium falciparum: two antigens of similar size are located in different compartments of the rhoptry. Exp Parasitol. 1990 70:193-206. Copyright Elsevier 2010.
See original on MMP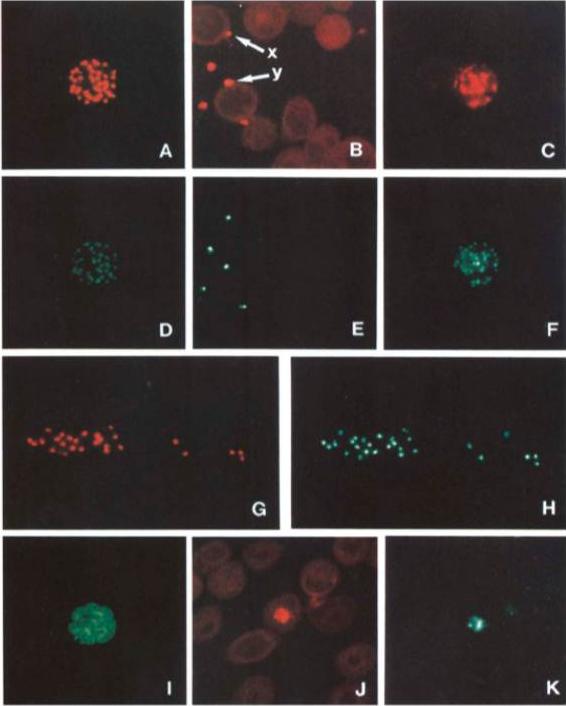
Single and dual immunofluorescent analysis of the subcellular localization of AMA-1, RAP-l and MSPl in mature schizonts, free merozoites, and ring stages of P. falciparum 7G8. Descriptors of the subcellular localization follow. Co-localized apical immunofluorescence of C-terminal AMA-1 and RAP-l in mature schizont, (A and D, respectively). Co-localized apical immunofluorescence of C-terminus-AMA-1 (x) and RAP-l in free merozoite, (B and E, respectively); surface localization of C-terminus-AMA-1 (y) and apical localization of RAP-l in free merozoite, (B and E, respectively). Co-localized apical immunofluorescence of N-termini-AMA-1 and RAP-l in mature schizont and free merozoite, (C, F and G, H, respectively). Surface localization of MSPl in mature schizont (I). Co-localization of C-terminus-AMA-1 and RAP-l in ring stage parasite, (J and K, respectively). Methanol-fixed schizont/ring stage thin films (ring development less than 3 h) were reacted with rat mAb IgG (25 pg ml-1), or rat mAb IgG and RAP-l culture supematant (dil. 15) in phosphate-buffered saline containing 1% foetal calf serum. Secondary stains were tetramethyl rhodamine isothiocyanate coupled or fluorescein isothiocyanate coupled anti-rat (A,B,C,G,J or I, respectively) and FITC coupled anti-mouse (human and rat adsorbed) (D,E,F,G and K).Narum DL, Thomas AW. Differential localization of full-length and processed forms of PF83/AMA-1 an apical membrane antigen of Plasmodium falciparum merozoites. Mol Biochem Parasitol. 1994 67:59-68. Copyright Elsevier 2009.
See original on MMP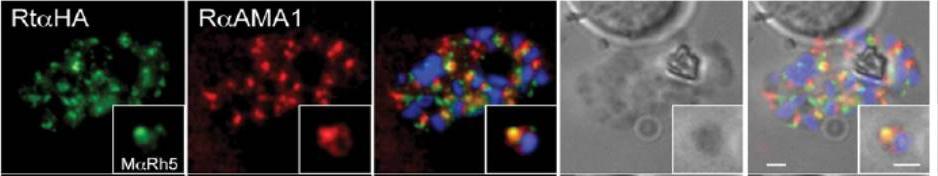
Immunofluorescence and phase contrast images of late schizonts or free merozoites (insets) to colocalise PfRh5 with AMA1. Each panel from left top right corresponds to anti-HA antibodies (to detect PhRh5), rabbit anti-AMA-1, overlay of each with DAPI nuclear stain, phase contrast and overlay of all images. Insets show individual merozoites. Scale bars = 1 mM. Baum J, Chen L, Healer J, Lopaticki S, Boyle M, Triglia T, Ehlgen F, Ralph SA, Beeson JG, Cowman AF. Reticulocyte-binding protein homologue 5 - An essential adhesin involved in invasion of human erythrocytes by Plasmodium falciparum. Int J Parasitol. 2008 39(3):371-80 Copyright Elsevier 2009.
See original on MMP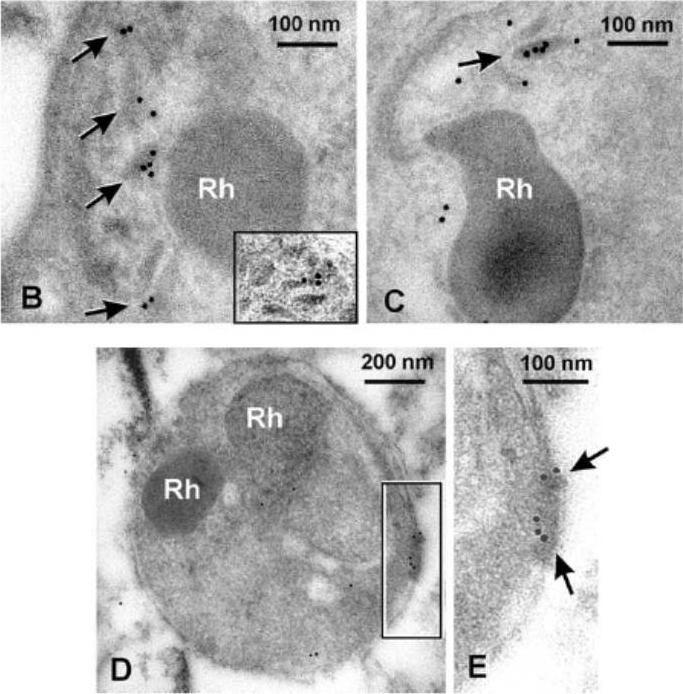
PfSUB2 Is a Microneme Protein. Electron micrographs showing immuno-gold labelling of micronemes within late-stage schizonts of PfSUB2HA clone 2D using: (B) anti-HA mAb 3F10, detecting epitope-tagged PfSUB2; and (C) a polyclonal antibody specific for PfAMA1. The inset in (B) shows another example of micronemal staining with mAb 3F10 from another schizont. Rh, rhoptry. (D and E) Posterior labelling of with anti-HA mAb 3F10 in a free merozoite of PfSUB2HA clone 2D. Arrows indicate immuno-gold labelling. Rh, rhoptry.Harris PK, Yeoh S, Dluzewski AR, O'Donnell RA, Withers-Martinez C, Hackett F, Bannister LH, Mitchell GH, Blackman MJ. Molecular identification of a malaria merozoite surface sheddase. PLoS Pathog. 2005 1(3):241-51.
See original on MMP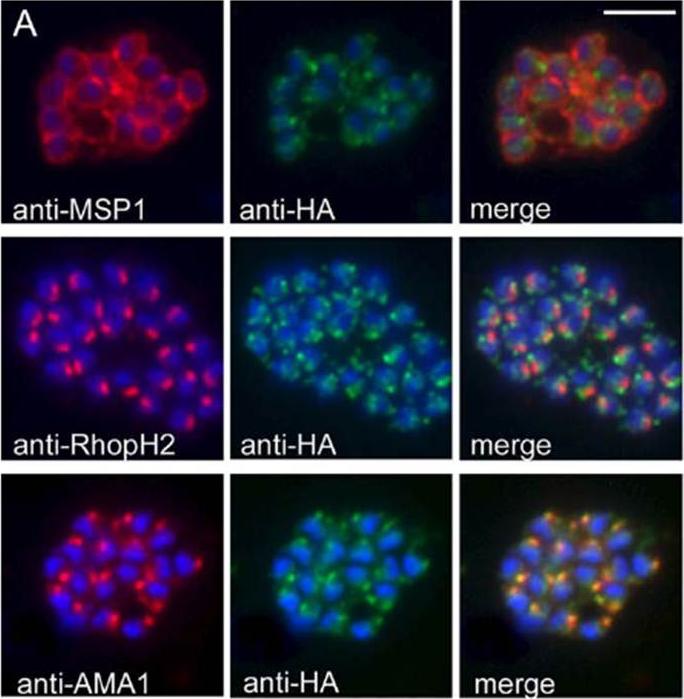
PfSUB2 Is a Microneme Protein. (A) IFA images of schizonts of PfSUB2HA clone 2D dual-labelled with mAbs X509 (anti-MSP1), 61.3 (anti-RhopH2), or 4G2 (anti-AMA1), plus in each case mAb 3F10 (anti-HA) The anti-HA signal co-localised only with the anti-AMA1 signal. Identical results were obtained with the uncloned transgenic PfSUB2HA line, and/or when anti-RAP2 mAb H5 was used as the rhoptry marker instead of mAb 61.3 (not shown). Parasite nuclei are stained throughout with DAPI (blue). Scale bar represents 2 lm.Harris PK, Yeoh S, Dluzewski AR, O'Donnell RA, Withers-Martinez C, Hackett F, Bannister LH, Mitchell GH, Blackman MJ. Molecular identification of a malaria merozoite surface sheddase. PLoS Pathog. 2005 1(3):241-51.
See original on MMP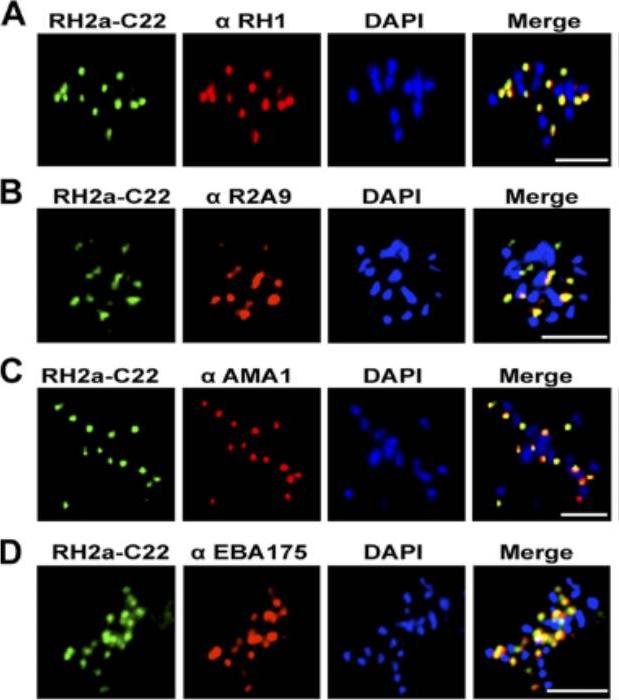
An immunofluorescence assay was performed on late-schizont-stage W2mef parasites stained for RH2a using the anti-RH2a MAb C22 and costained either with an antibody against the rhoptry neck marker RH1 (A), rabbit polyclonal antibody R2A9, recognizing both RH2a and RH2b (B), or an antibody against the micronemal marker AMA1 (C) or EBA175 (D). Bars, 5 mm. PfRH2a showed a punctate pattern at the apical tip of the merozoite in schizonts and appeared to colocalize with PfRH1 in W2mef and 3D7 parasites, strongly suggesting that RH2a is located at the rhoptry neck.Gunalan K, Gao X, Liew KJ, Preiser PR. Differences in erythrocyte receptor specificity of different parts of the Plasmodium falciparum reticulocyte binding protein homologue 2a. Infect Immun. 2011 79:3421-30.
See original on MMP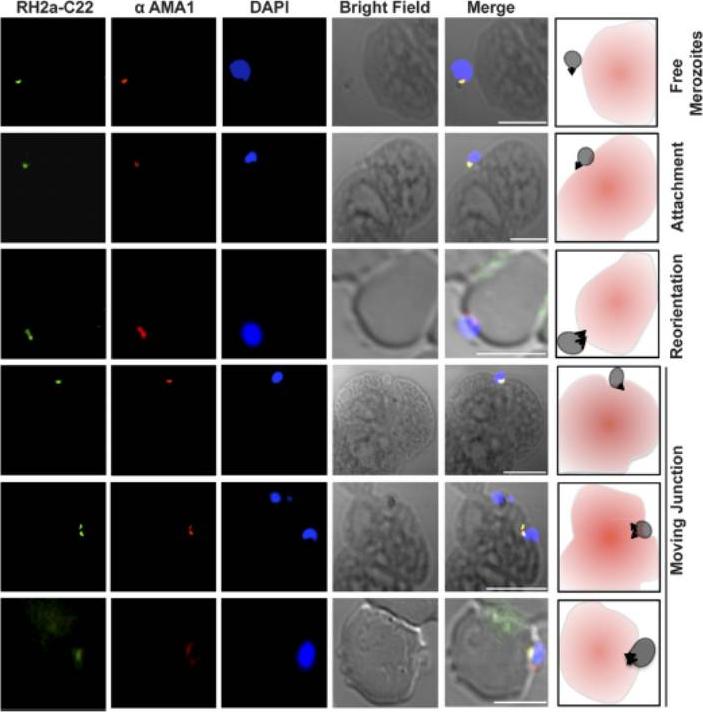
RH2a follows the moving junction during invasion. RH2a was costained with AMA1. From top to bottom, immunofluorescence images show merozoites progressing through different stages of invasion: free merozoites, an attached merozoite, a reoriented merozoite whose apical end is facing the red blood cells, and the progression of the moving junction. Bars, 5 mm.Gunalan K, Gao X, Liew KJ, Preiser PR. Differences in erythrocyte receptor specificity of different parts of the Plasmodium falciparum reticulocyte binding protein homologue 2a. Infect Immun. 2011 79:3421-30. PMID:
See original on MMP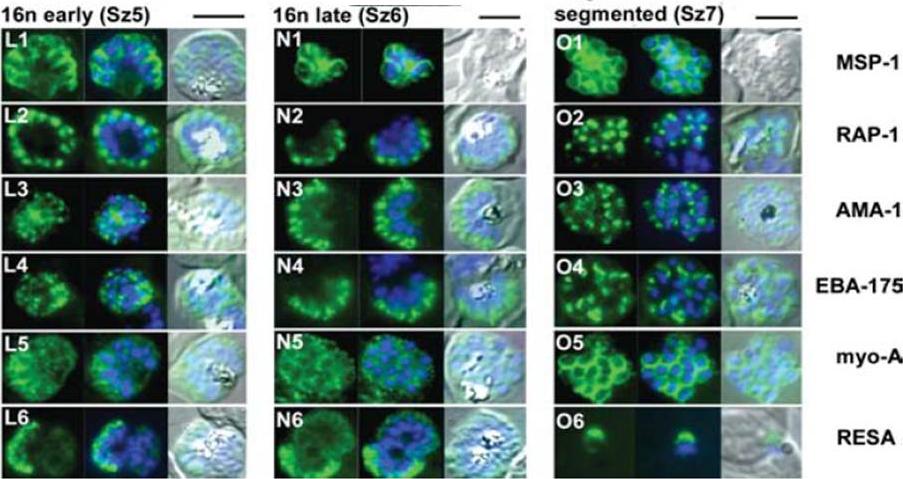
The appearance of Plasmodium falciparum schizonts of the ITO4 line at different times of development. The time of sampling from invasion and the number of nuclei per schizont are indicated. Antibodies to proteins associated with invasion related organelles or the parasite periphery (rhoptries : RAP-1; micronemes: AMA-1, EBA-175; dense granules: RESA; parasite periphery: MSP-1, myo-A) were chosen to follow the development and assembly of merozoites in maturing schizonts.Margos G, Bannister LH, Dluzewski AR, Hopkins J, Williams IT, Mitchell GH. Correlation of structural development and differential expression of invasion-related molecules in schizonts of Plasmodium falciparum. Parasitology. 2004 129:273-87. Copyright Cambridge University Press Journals 2011
See original on MMP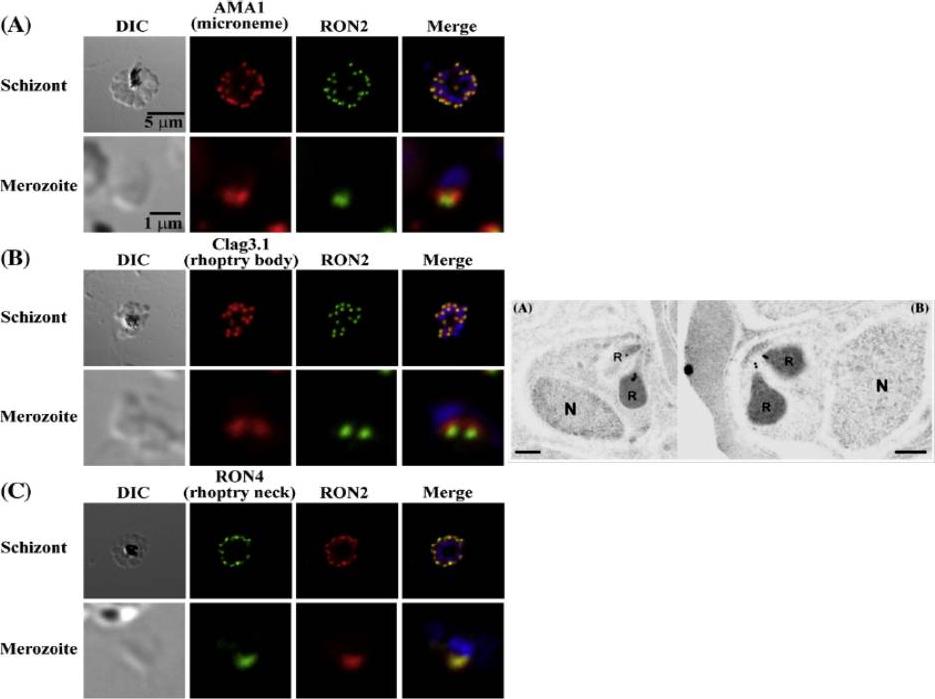
Left: PfRON2 is expressed at the apical end of Plasmodium merozoites. Schizont infected erythrocytes and merozoites were dual-labeled with antisera against PfRON2 and PfAMA1 PF11_0344 (A), PfClag3.1 PFC0110w (B), or PfRON4 PF11_0168 (C). Merged images are shown in the right panels. All segmented schizonts and merozoites are positive for PfRON2. Nuclei are counterstained with DAPI. Colocalization of PfRON2 with PfRON4 (rhoptry neck marker) was observed but neither colocalized with PfClag3.1 (rhoptry body marker) nor PfAMA1 (microneme marker). Right: Longitudinally sectioned merozoites in schizont-infected erythrocytes were labeled with anti-PfRON2 serum followed by secondary Ab conjugated with gold particles. Gold particles were restricted to the narrow neck portion of the rhoptries (R). Two different images are shown (A and B). N indicates nucleus. Bars=200 nm. Cao J, Kaneko O, Thongkukiatkul A, Tachibana M, Otsuki H, Gao Q, Tsuboi T, Torii M. Rhoptry neck protein RON2 forms a complex with microneme protein AMA1 in Plasmodium falciparum merozoites. Parasitol Int. 2009 58:29-35.
See original on MMP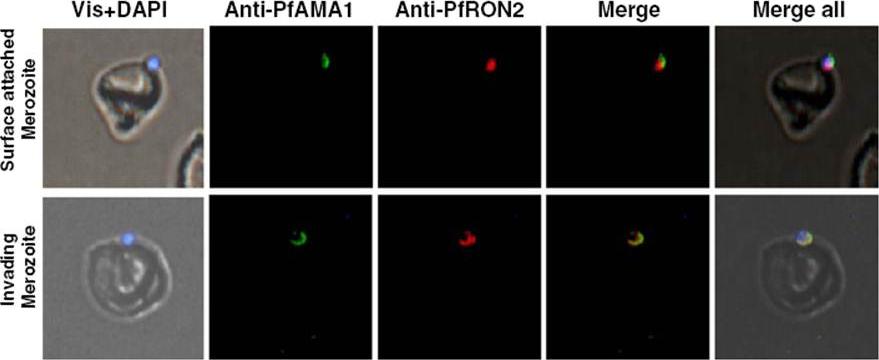
Immunofluorescence assay showing co-localization of PfRON2 with PfAMA1 at apical end at the time of meozoite attachment during merozoite invasion. At the time of attachment of the merozoite with the host RBCs, the PfRON2 staining was at the apical tip, the PfAMA1 staining was also mainly at the apical tip and with some distribution at the merozotie surface. During parasite invasion, the PfRON2 and PfAMA1 showed col-ocalization at the apical end of merozoite.Hossain ME, Dhawan S, Mohmmed A. The cysteine-rich regions of Plasmodium falciparum RON2 bind with host erythrocyte and AMA1 during merozoite invasion. Parasitol Res. 2011 110(5):1711-21
See original on MMP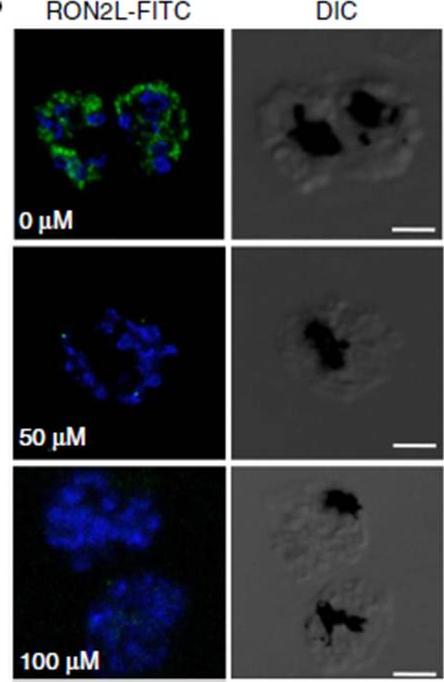
Immunofluorescence assay using FITC-labelled RON2L peptide. FITC-labelled RON2 peptide binds to AMA1 (blue)in the mature schizonts in the absence of inhibitors, whereas pre-incubation with inhibitor NCGC00015280 prevents binding of the peptide. the binding of RON2 to AMA1 triggers junction formation, which commits the merozoite for invasion. Although FITC-labelledPfRON2L binds to schizonts, binding is prevented in the presence of the inhibitor.Srinivasan P, Yasgar A, Luci DK, Beatty WL, Hu X, Andersen J, Narum DL, Moch JK, Sun H, Haynes JD, Maloney DJ, Jadhav A, Simeonov A, Miller LH. Disrupting malaria parasite AMA1-RON2 interaction with a small molecule prevents erythrocyte invasion. Nat Commun. 2013 4:2261.
See original on MMP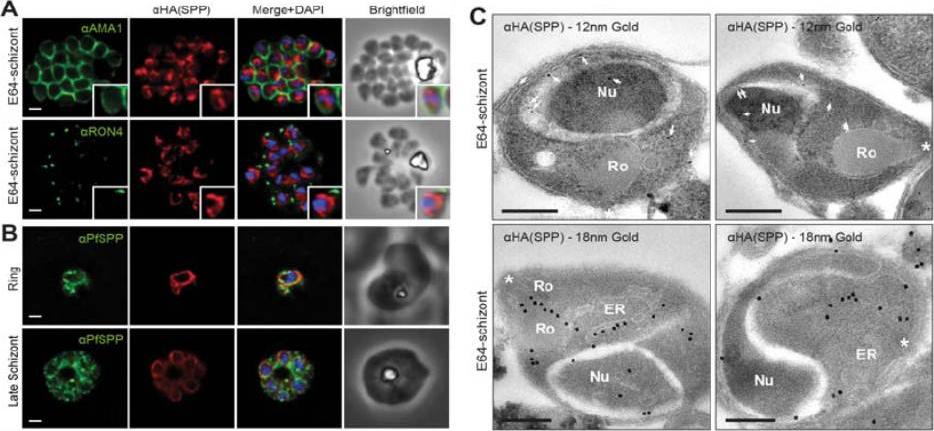
PfSPP localises to the parasite nuclear periphery.(A) IFA of E64 treated PfSPP-HA schizonts labeled with anti-PfAMA1, PfRON4 and HA antibodies. Scale bar 1 μm. Inset displays magnification of single merozoite. (B) IFA of ring and late schizont stage PfSPP-HA parasites labeled with anti-PfSPP and HA antibodies. Scale bar 1 mm. (C) Immuno-electron micrograph of E64 treated PfSPP-HA schizonts with 12 nm or 18 nm immunogold. Arrowheads mark 12 nm immunogold labels and white asterisks marks parasite apex. Nucleus (Nu), endoplasmic reticulum (ER) and rhoptry (Ro). Scale bar = 200 nm. PfSPP is an ER-resident peptidase that remains intracellular throughout the invasion process.Marapana DS, Wilson DW, Zuccala ES, Dekiwadia CD, Beeson JG, Ralph SA, Baum J. Malaria parasite signal peptide peptidase is an ER-resident protease required for growth but not invasion. Traffic. 2012 13(11):1457-65
See original on MMP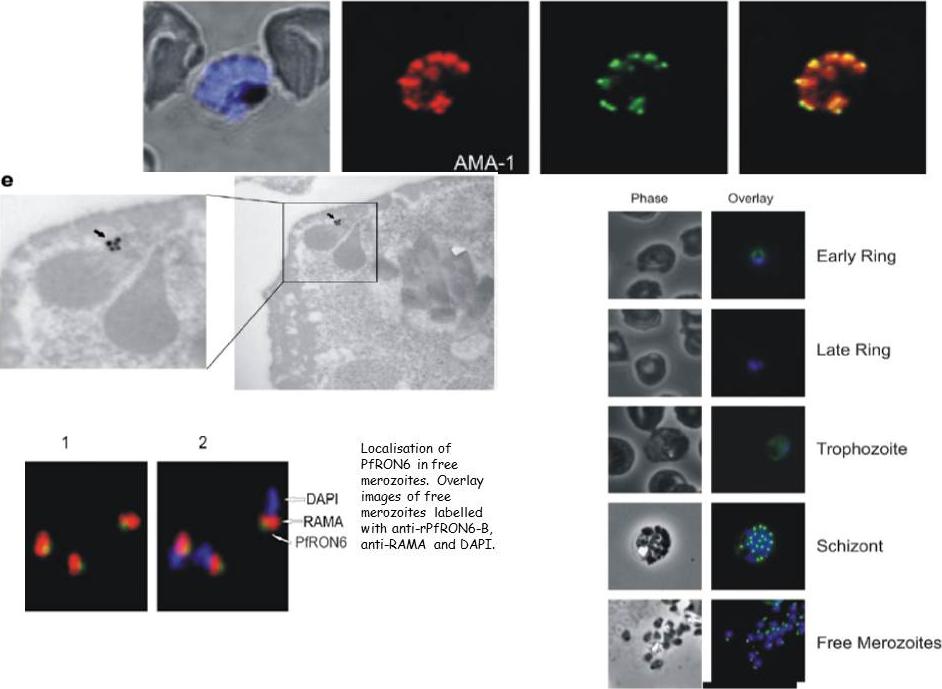
Upper panel: Double labelling of PfRON6 with AMA1 PF11_0344 in segmented schizonts. Confocal microscopy using anti-rPfRON6-B with anti-AMA1 (microneme marker). EM: Electron micrograph of immunogold staining using affinity purified anti-rRON6-C antibodies. Arrows indicate PfRON6.Right panel: Localisation of PfRON6 during the asexual red blood cell (RBC) cycle. Immunofluorescence microscopy using anti-rPfRON6-B antisera and DAPI on parasites at different stages of the asexual RBC cycle. The corresponding phase contrast images are shown.Proellocks NI, Kats LM, Sheffield DA, Hanssen E, Black CG, Waller KL, Coppel RL. Characterisation of PfRON6, a Plasmodium falciparum rhoptry neck protein with a novel cysteine-rich domain. Int J Parasitol. 2008 39(6):683-92. Copyright Elsevier 2009.
See original on MMP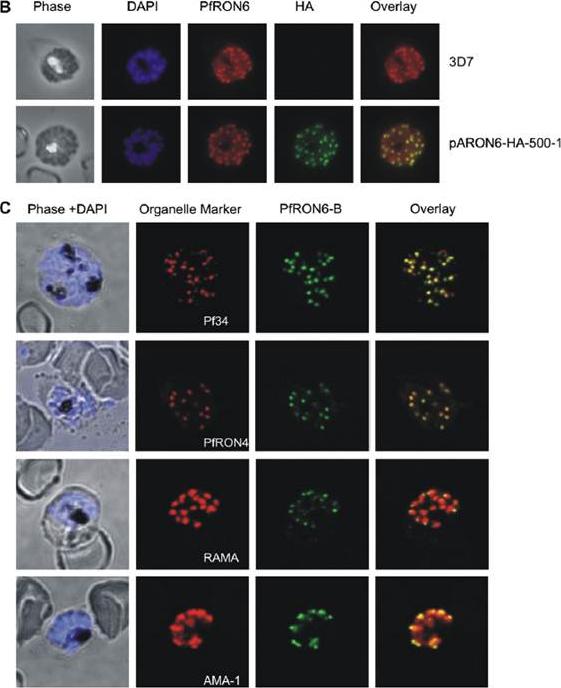
Localisation of the PfRON6 protein in parasitised red blood cells. (B) Double labelling using anti-rPfRON6-B and anti- haemagglutinin (HA) in HA-tagged transgenic parasites. Confocal microscopy using anti-rPfRON6-B with anti-HA. Corresponding overlay images are shown. (C) Double labelling of PfRON6 with Pf34, PfRON4, RAMA or AMA1 in segmented schizonts. Confocal microscopy using anti-rPfRON6-B with anti-Pf34 (rhoptry neck marker), anti-PfRON4 (rhoptry neck marker), anti-RAMA (rhoptry bulb marker), or anti-AMA1 (microneme marker). Corresponding overlay images are shown. a punctate pattern of fluorescence in segmented schizonts characteristic of localisation with the apical secretory organelles.Proellocks NI, Kats LM, Sheffield DA, Hanssen E, Black CG, Waller KL, Coppel RL. Characterisation of PfRON6, a Plasmodium falciparum rhoptry neck protein with a novel cysteine-rich domain. Int J Parasitol. 2008 39(6):683-92. Copyright Elsevier 2009.
See original on MMP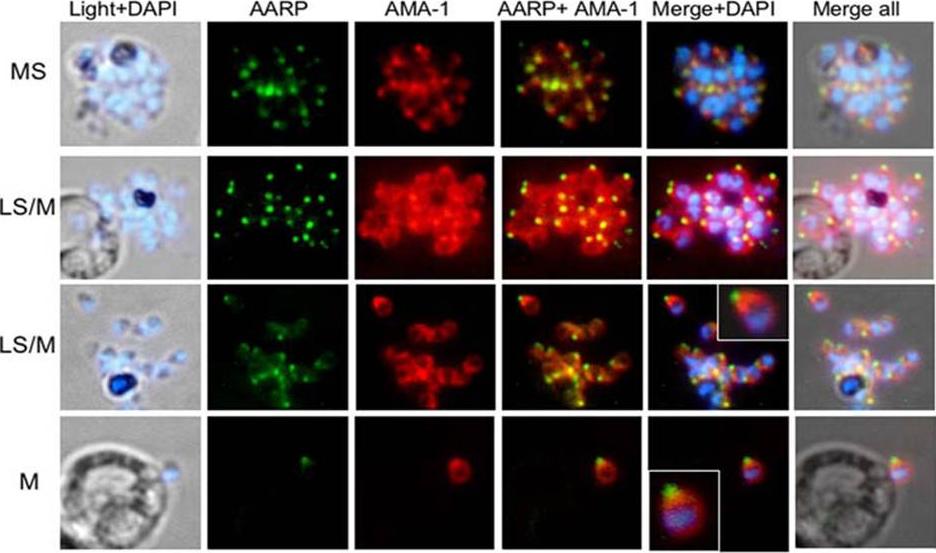
Spatial localization of merozoite’s microneme resident proteins AMA-1 and PfAARP by co-immunostaining with anti-PfAARP (green) and anti-AMA-1 (red) Abs: both showed punctate staining in the schizonts. In the late schizonts and merozoites, AMA-1 was present over the entire surface of the merozoites but is most densely distributed at their apical tip. Enlarged image of selected individual merozoite is shown in the inset. Wickramarachchi T, Devi YS, Mohmmed A, Chauhan VS. Identification and characterization of a novel Plasmodium falciparum merozoite apical protein involved in erythrocyte binding and invasion. PLoS ONE. 2008 Mar 5;3(3):e1732.
See original on MMP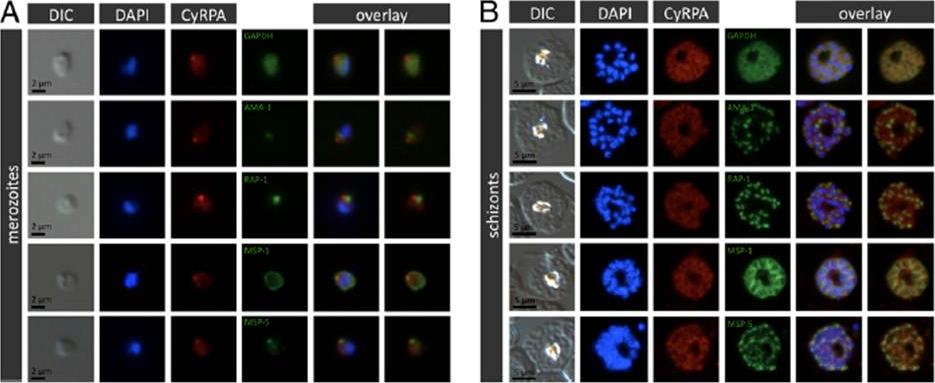
Localization of CyRPA to the merozoite apex by immunofluorescence staining. P. falciparum 3D7 merozoites (A) or schizont stages (B) were coimmunostained with anti-CyRPA mAb c06 (red) and anti-GAPDH Abs (marker for cytosol), AMA-1 (marker for micronemes), RAP-1 (marker for rhoptry bulbs), MSP-1 (marker for merozoite surface), or MSP-5 (green). Nuclei were stained with DAPI (blue). Original magnification x1008. CyRPA is localized at the merozoite apex.Dreyer AM, Matile H, Papastogiannidis P, Kamber J, Favuzza P, Voss TS, Wittlin S, Pluschke G. Passive immunoprotection of Plasmodium falciparum-infected mice designates the CyRPA as candidate malaria vaccine antigen. J Immunol. 2012 188(12):6225-37.
See original on MMP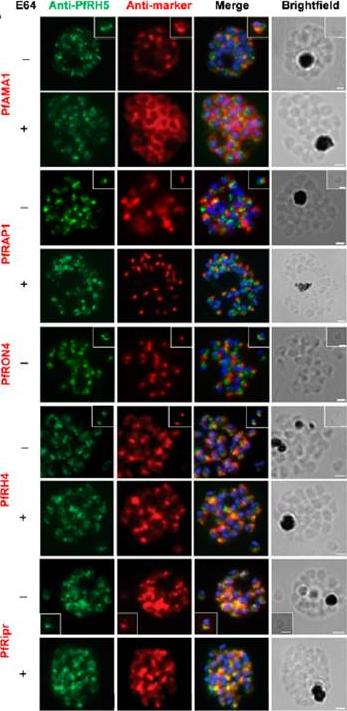
Localization of PfRH5 was assessed by indirect IFA using anti-PfRH5 polyclonal rabbit serum (green). Fixed and permeabilized schizonts with (+) or without (2) E64 treatment or free merozoites (inset) of 3D7 clone P. falciparum were costained with mouse Abs (red) to mark various organelles: PfAMA1 polyclonal (microneme), PfRAP1 mAb (rhoptry body), or PfRON4 mAb (rhoptry neck); or polyclonal mouse serum against further Ags: PfRH4 and PfRipr. Figures show the merge of the dual staining Abs and nuclei stained with DAPI (blue), as well as the brightfield view. Scale bars, 1 mm. anti-PfRH5 rabbit Abs (8) do not colocalize with conventional markers of the rhoptry bulb, rhoptry neck, or micronemes (PfRAP1, PfRON4, and PfAMA1, respectively) in permeabilized schizonts or free merozoites of 3D7 clone P. falciparum parasites. Minimal colocalization in schizonts with PfRH2a/b (data not shown) and also with PfRH4, described as a marker of the rhoptry tip, but significant colocalization with PfRipr in merozoites and late-stage schizonts, especially following treatment with the cysteine protease inhibitor E64, which prevents merozoite release by inhibiting schizont rupture. Impossible to detect PfRH5 on the merozoite surface in any assay, with staining only successful following permeabilization .Douglas AD, Williams AR, Knuepfer E, Illingworth JJ, Furze JM, Crosnier C, Choudhary P, Bustamante LY, Zakutansky SE, Awuah DK, Alanine DG, Theron M, Worth A, Shimkets R, Rayner JC, Holder AA, Wright GJ, Draper SJ. Neutralization of Plasmodium falciparum Merozoites by Antibodies against PfRH5. J Immunol. 2014 192(1):245-58.
See original on MMP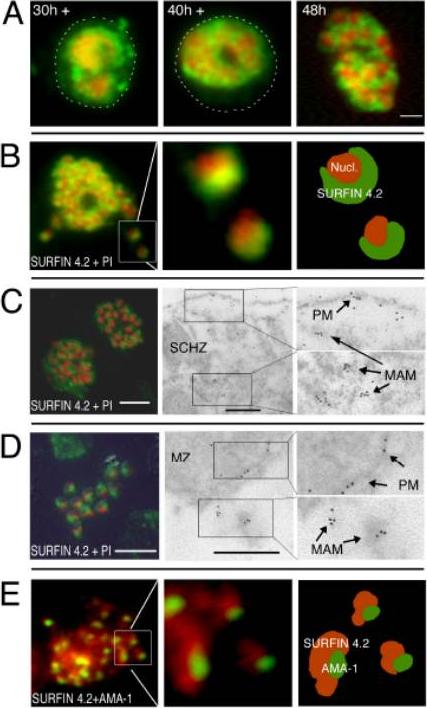
SURFIN accumulates within the parasitophorous vacuole in late schizonts and is associated with released merozoites. (A) Alexa 488–labeled SURFIN4.2 (green) detected with anti-S1.3 antibodies in parasites developing from late trophozoite stage to schizonts. PI staining (red) was used to visualize nuclei. (B–D) SURFIN4.2 was found localized in MAM and the PM of late and bursting schizonts (48 h), and associated with the released merozoite surface. Dashed circles indicate the area of the IE. (B) IFA with anti-S1.3 + PI on bursting schizonts. IFA (anti-rSURFIN4.2) and immunoelectron microscopy (anti-S1.4 Ig) on late schizonts (C) and released merozoites (D). SURFIN associated with PM or merozoite-associated material is indicated by arrows. (E) SURFIN4.2 did not colocalize with the microneme AMA-1. A detailed analysis of SURFIN4.2 (anti-S1.3 antibodies) and AMA-1 in relation to the nuclei and to each other in merozoites is shown. (B and E) A schematic representation of the antigen localization is shown on the right. Bars: IFA, 2 mm; EM, 0.5 mm.Winter G, Kawai S, Haeggström M, Kaneko O, von Euler A, Kawazu S, Palm D, Fernandez V, Wahlgren M. SURFIN is a polymorphic antigen expressed on Plasmodium falciparum merozoites and infected erythrocytes. J Exp Med. 2005 201:1853-63.
See original on MMP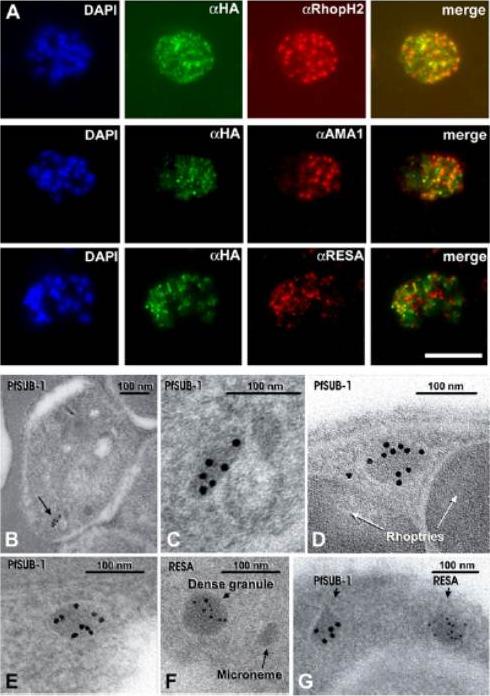
(A) Schizonts of 3D7SUB1HA3 clone C10 dual-labeled with the anti-HA mAb 3F10 (aHA; green) plus mAb 61.3 (aRhopH2), mAb 4G2 (aPfAMA1; micronemes, or mAb 28/2 (aRESA; dense granules. Nuclei were stained with DAPI (blue). Merged images (no DAPI) show that PfSUB1 localizes with none of the other markers. The scale bar represents 5 mm. Identical results were obtained with 3D7SUB1HA3 clone F7 or by using mAbs specific for RAP2 or EBA-175 as markers for rhoptries and micronemes respectively (data not shown). (B–G) Immunoelectron microscopic localization of PfSUB1 in P. falciparum schizonts. (B and C) Labeling of an elongated organelle, arrowed in (B) and at higher magnification in (C), by anti-PfSUB1 antibodies labeled with 10 nm immunogold. (D and E) Two more examples of PfSUB1 positive organelles are shown, also labeled with 10 nm immunogold. These are less elongate than in (B) and (C), reflecting some variation in organelle shape; however, they are typically ellipsoidal, in contrast with the rounded dense granules. (F and G) Staining for RESA with mAb 28/2 and 5 nm immunogold. In (F), an unlabeled microneme is also shown for size comparison. In (G) double staining for PfSUB1 and RESA with different sizes of immunogold shows that the two proteins are in different organelles.Yeoh S, O'Donnell RA, Koussis K, Dluzewski AR, Ansell KH, Osborne SA, Hackett F, Withers-Martinez C, Mitchell GH, Bannister LH, Bryans JS, Kettleborough CA, Blackman MJ. Subcellular discharge of a serine protease mediates release of invasive malaria parasites from host erythrocytes. Cell. 2007 131:1072-83.
See original on MMP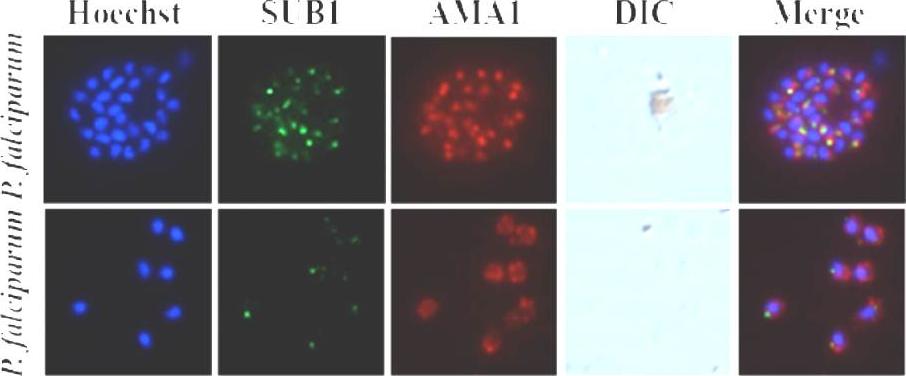
Indirect immunofluorescence assays on air-dried P.falciparum 3D7 segmented schizonts (upper), individual merozoites (lower). Merozoite nuclei are labeled with Hoescht 33342. The scale bar represents 3μm. A typical punctate pattern is seen with partial superposition with the signal produced by the anti-AMA1 mAb 28G2. This is in line with the subcellular localization of PfSUB1 and AMA1 in the exonemes and micronemes, respectively. Bouillon A, Giganti D, Benedet C, Gorgette O, Pêtres S, Crublet E, Girard-Blanc C, Witkowski B, Ménard D, Nilges M, Mercereau-Puijalon O, Stoven V, Barale JC. In silico screening on the 3D-model of the Plasmodium vivax SUB1 protease leads to the validation of a novel anti-parasite compound. J Biol Chem. 2013 288(25):18561-73
See original on MMP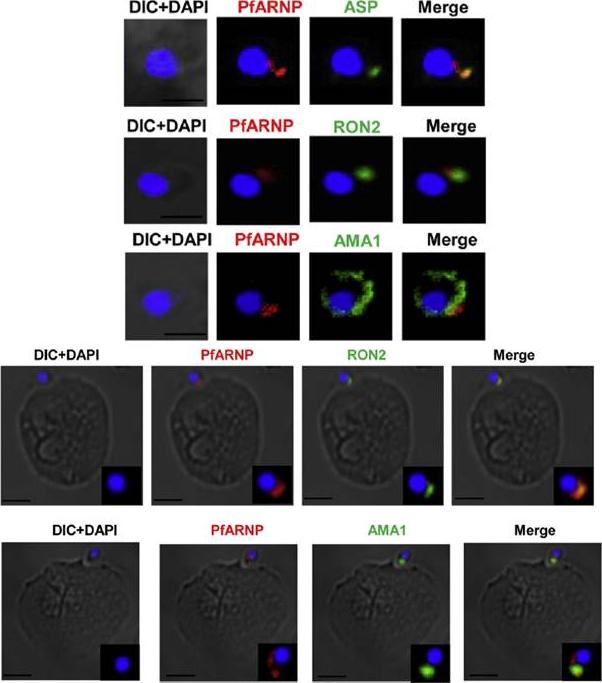
Localization of PfARNP in free merozites and at the tight junction. (Upper panel) IFA of merozoite using anti PfARNP antibody. Co-staining of PfARNP (red) was performed with ASP, RON2 (1973–2067 aa) and AMA1 (green) in free merozoites. Staining was observed at the apex of merozoite distinct from nuclear stain DAPI (blue). The staining merged perfectly well with ASP, RON2 and not with AMA1 confirming the localization of PfARNP in neck of rhoptries of merozoites. Scale bar shows 1 mm. (Lower panel) Localization of PfARNP during invasion of merozoite. IFA of PfARNP was performed with RON2 and AMA1, known markers of tight junction in Cytochalasin D treated merozoites invading to erythrocytes. The labeling of PfARNP (red) was observed at the site of attachment of merozoite to erythrocyte. The staining of PfARNP merged perfectly with RON2 and AMA1(green), indicating that PfARNP is present at the site of tight junction. The inset of each image show the optically zoomed view of invading merozoite. Scale bar shows 2 mm. Hans N, Singh S, Jain SK, Chauhan VS. Identification of novel rhoptry neck protein of Plasmodium falciparum. Mol Biochem Parasitol. 2013 188(1):34-9 Copyright Elsevier
See original on MMP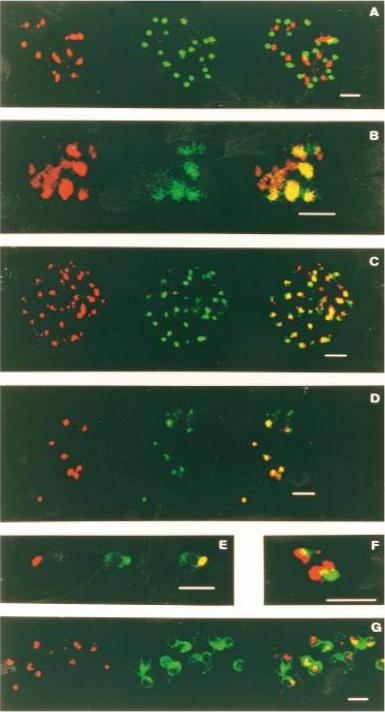
Localization and co-localization of PfPK4, RAP-1, PF14_0102, PF83/AMA-1 PF11_0344 and MSP1 PFI1475w. (A) A segmenter, stained for PfPK4 (red) and RAP-1 (green), with a composite of both labels on the right ; (B) schizont ; (C) segmenter ; (D) and (E) free merozoites, stained for PfPK4 (red) and PF83/AMA-1 (green), with composites on the right ; (F) a free merozoite, a composite of PfPK4 (red) and RAP-1 (green); (G) free merozoites stained for PfPK4 (red) and MSP-1 (green), with a composite of both labels on the right. The scale bars represent 2 mm. PfPK4 distributed throughout the trophozoite, whereas it was found in discrete foci (punctate distribution) in segmenters. PfPK4 co-localizes with AMA-1 at the apical complex in segmenters and merozoites, but does not co-localize with RAP-1.Reproduced with permission, from Möhrle JJ, Zhao Y, Wernli B, Franklin RM, Kappes B. Molecular cloning, characterization and localization of PfPK4, an eIF-2a kinase-related enzyme from the malarial parasite Plasmodium falciparum. Biochem J. 1997 328:677-87. © the Biochemical Society.
See original on MMP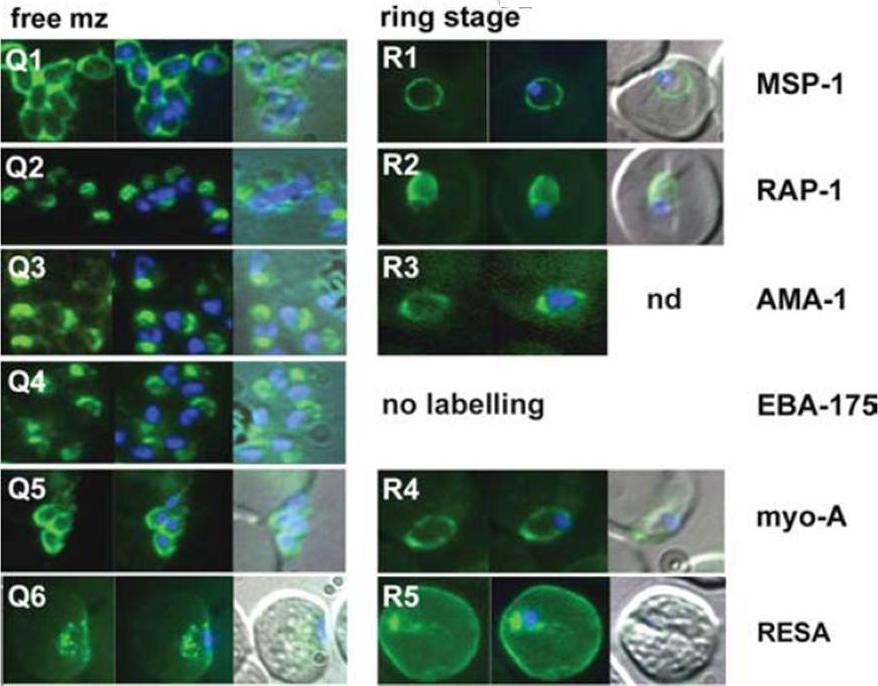
The appearance of Plasmodium falciparum merozoites and rings of the ITO4 line at different times of development. Antibodies to proteins associated with invasion related organelles or the parasite periphery (rhoptries : RAP-1; micronemes: AMA-1, EBA-175; dense granules: RESA; parasite periphery: MSP-1, myo-A) were chosen to follow the development and assembly of merozoites and rings.Margos G, Bannister LH, Dluzewski AR, Hopkins J, Williams IT, Mitchell GH. Correlation of structural development and differential expression of invasion-related molecules in schizonts of Plasmodium falciparum. Parasitology. 2004 129:273-87. Copyright Cambridge University Press Journals 2011
See original on MMP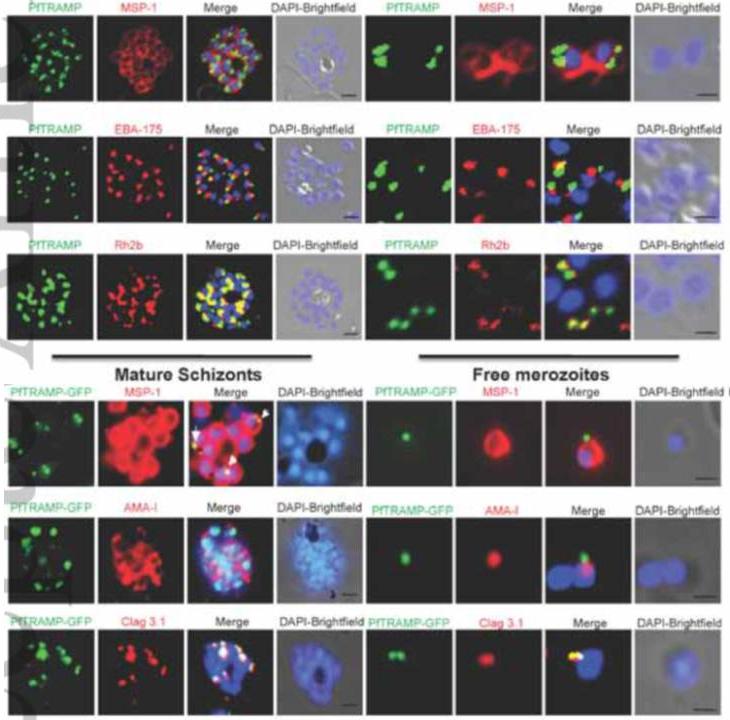
PfTRAMP localizes to the rhoptries of P. falciparum schizonts and merozoites.(A) Co-localization of PfTRAMP with MSP1, EBA175 and Rh2b in schizonts and free merozoites by IFA. PfTRAMP (green) co-localizes with rhoptry marker PfRh2b (red) but does not co-localize with EBA175 (red) or merozoite surface marker MSP1 (red) in schizonts and merozoites. (B)Co-localization of PfTRAMP-GFP in transgenic P. falciparum 3D7 with MSP1, AMA1 and Clag3.1. PfTRAMP-GFP (green) co-localizes with rhoptry marker Clag 3.1 (red) but not with microneme markerPfAMA1 (red) or merozoite suface marker PfMSP1 (red) in schizonts and merozoites. Nuclei were stained with DNA intercalating dye DAPI.Siddiqui FA, Dhawan S, Singh S, Singh B, Gupta P, Pandey A, Mohmmed A, Gaur D, Chitnis CE. A Thrombospondin Structural Repeat Containing Rhoptry Protein from Plasmodium falciparum Mediates Erythrocyte Invasion. Cell Microbiol. 2013 15(8):1341-56
See original on MMP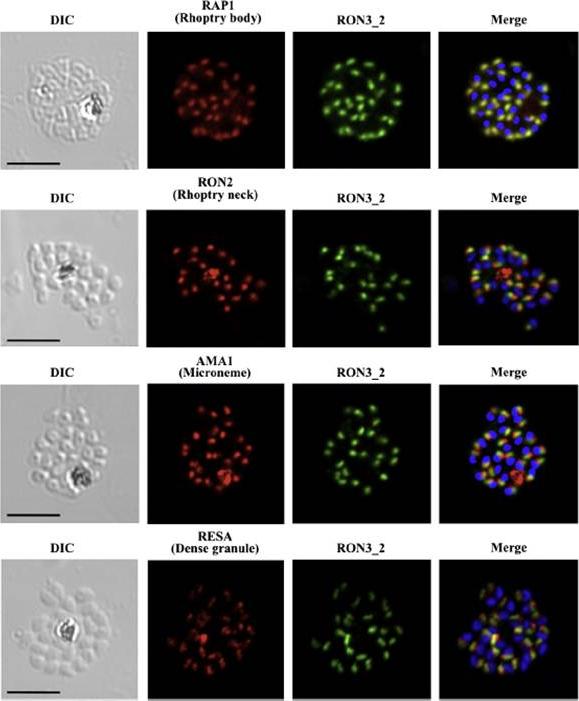
PfRON3 is expressed at the apical end of Plasmodium merozoites. Schizont and merozoite stage parasites were dual-labeled with antisera against PfRON3_2 and either PfRAP1 (rhoptry body marker), or PfRON2 (rhoptry neck marker), or PfAMA1 (microneme marker), or PfRESA (dense granule marker). Nuclei are visualized with DAPI in merged images shown in the right panels. Bars represent 5 μm. PfRON3 is localized to the but in the rhoptry body.Ito D, Han ET, Takeo S, Thongkukiatkul A, Otsuki H, Torii M, Tsuboi T. Plasmodial ortholog of Toxoplasma gondii rhoptry neck protein 3 is localized to the rhoptry body. Parasitol Int. 2011 60(2):132-8.
See original on MMP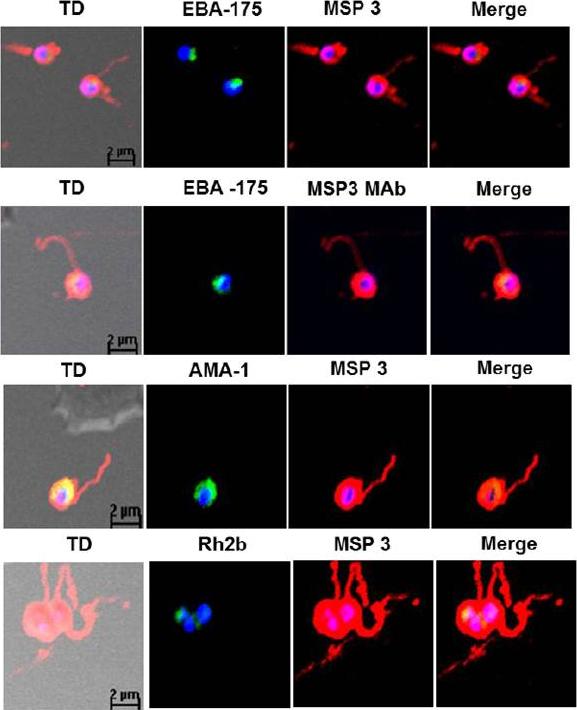
Detection of fibrillar structures in Plasmodium falciparum merozoites by immunofluorescence assay. Co-immunostaining of free merozoites with anti-MSP3 Abs along with Abs against EBA-175, AMA-1, Rh2b and MSP5. In all the slides long fibrous structure attached to free merozoite was detected by anti-MSP3 Abs. Monoclonal MSP3 antibodies also stained these fibrillar structures associated with merozoites, but other marker protein did not detect fibrillar assemblies. The long fibrils like structures attached at one end with merozoite surface were consistently seen to associate to distant erythrocytes as well as wrap around them.Imam M, Singh S, Kaushik NK, Chauhan VS. Plasmodium falciparum Merozoite Surface Protein 3: oligomerization, self-assembly and heme complex formation. J Biol Chem. 2013 289(7):3856-68
See original on MMP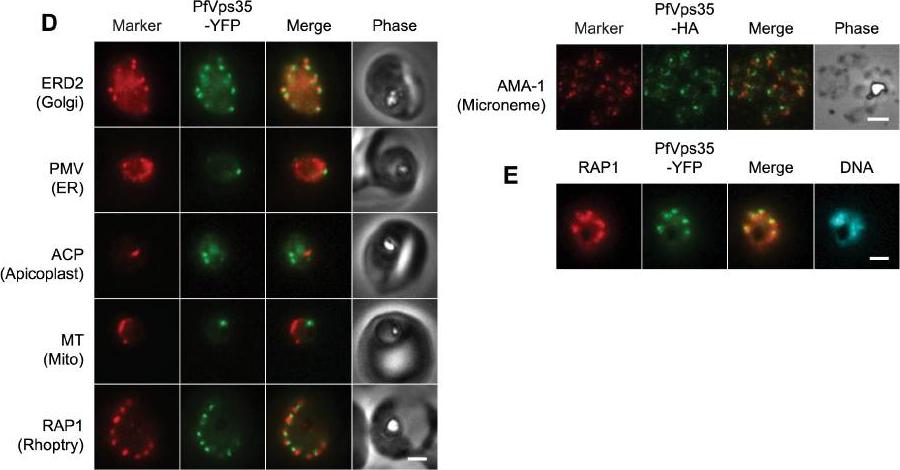
(D) Co-localization of PfVps35-YFP or PfVps35-HA and organellar markers in fixed parasites (except for MitoTracker, which was imaged live). PMV, plasmepsin V; ACP, acyl carrier protein; MT, MitoTracker Red CM-H2Xros; RAP1, rhoptry associated protein 1; AMA1, apical membrane antigen 1. The AMA1 panel shows free merozoites; all others are intraerythrocytic. Organelles labeled by the markers are indicated in parenthesis. Marker-derived fluorescence is pseudocolored red. (E) PfVps35-YFP is adjacent to developing rhoptries in a 2N parasite. Hoechst 33342 fluorescence (DNA) is pseudocolored cyan. In all panels, YFP fluorescence is pseudocolored green. Scale bars, 2 mm.Krai P, Dalal S, Klemba M. Evidence for a Golgi-to-Endosome Protein Sorting Pathway in Plasmodium falciparum. PLoS One. 2014 9(2):e89771.
See original on MMP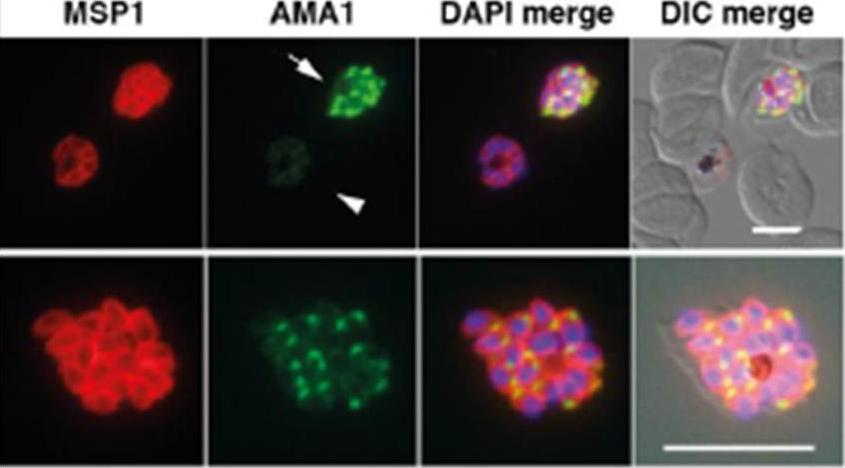
Schizonts of rapamycin-treated parasites were dual labelled with anti-MSP1 (red) and anti-PfAMA1 (green) to show reduction in PfAMA1 expression. Schizonts displaying a gradient of high (arrow, top row), medium (bottom row) and low (arrowhead, top row) levels of PfAMA1 expression were observed. Scale bar is 5 μm. All of the merozoite microneme organelles within any particular parasite cell were evenly stained indicating that the extent of PfAMA1 knockdown was uniform between daughter merozoites.Yap A, Azevedo MF, Gilson PR, Weiss GE, O'Neill MT, Wilson DW, Crabb BS, Cowman AF. Conditional expression of apical membrane antigen 1 in Plasmodium falciparum shows it is required for erythrocyte invasion by merozoites. Cell Microbiol. 2014 Feb 27. [Epub ahead of print]
See original on MMP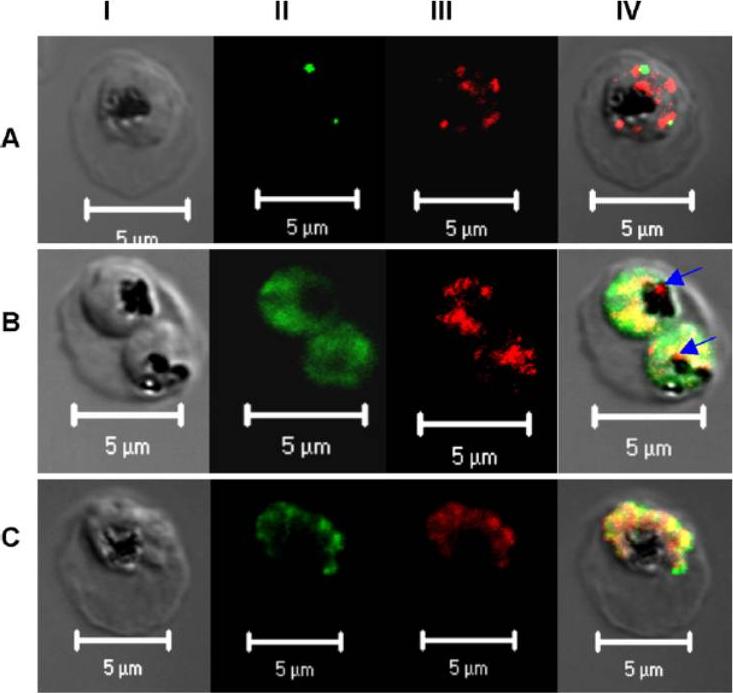
Subcellular localization of PfPRL. Micrographs represent the co-staining profiles of PfPRL with the Golgi marker PfErd2 (A), the ER marker PfBip (B), and the rhoptry/micronemal protein PfAMA-1 (C). Synchronous 3D7 parasite cultures were probed with purified rabbit anti-PfPRL in combination with antibodies to the compartmental markers. Binding of primary antibodies was detected using Alexa-Fluor 488 conjugated antibodies (organellar markers) and Alexa-Fluor 555-conjugated anti-rabbit IgG (PfPRL). Panel I is differential interference contrast image, panel II is fluorescence due to Alexa-Fluor 488 (green), panel III fluorescence due to Alexa-Fluor 555 (red), while panel IV is a merge of the first three panels. Yellow spots indicate overlap between PfPRL and the respective organellar marker, while blue arrows indicate food PfPRL-associated sites in the digestive vacuole.Pendyala PR, Ayong L, Eatrides J, Schreiber M, Pham C, Chakrabarti R, Fidock DA, Allen CM, Chakrabarti D. Characterization of a PRL protein tyrosine phosphatase from Plasmodium falciparum. Mol Biochem Parasitol. 2008 158(1):1-10.
See original on MMP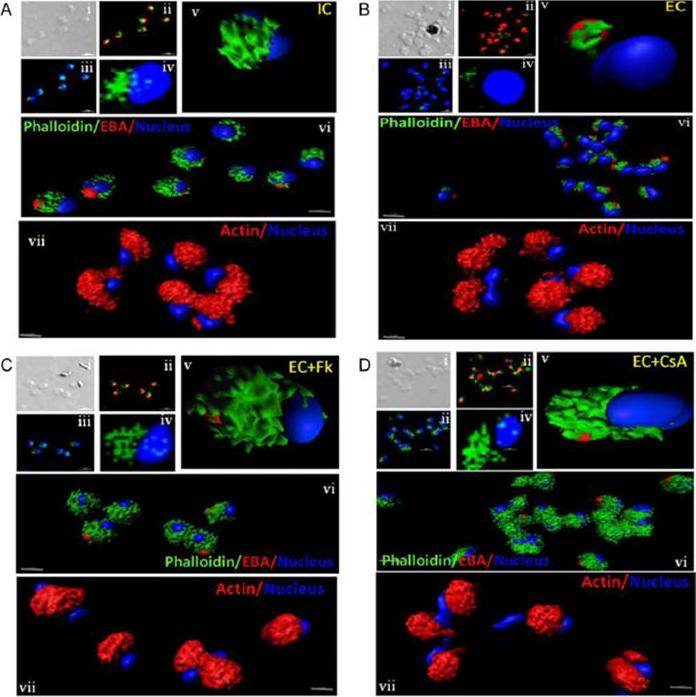
Actin polymerization status in P. falciparum merozoites. Merozoites were stained with Alexa 488-labeled phalloidin to detect polymerized F-actin (green), mouse anti EBA-175 mouse sera and Alexa Fluor 594-conjugated anti-mouse IgG goat antibodies to define location of micronemes (red) and DAPI to define location of nuclei (blue). Slides were observed under a confocal laser microscope (Nikon A1R). Bright field (i) and single slice confocal fluorescence images of merozoites (ii – iv) are shown. Three-dimensional reconstruction of confocal z stack fluorescence images of merozoites was performed using Imaris software (v –vii). Total actin (polymerized F-actin and monomeric G-actin) was detected with anti-actin rabbit sera followed by Alexa Fluor 594-conjugated anti-rabbit IgG goat antibodies (red in vii). Levels of total actin are similar in all conditions tested. Transfer of merozoites from IC to EC buffer leads to disassembly of polymerized F-actin at the apical tip of merozoites. Treatment of merozoites with FK506 and CsA prior to transfer to EC buffer leads to accumulation of polymerized actin at the apical end. Actin depolymerizing agent CytD and actin stabilizing agent JAS were used as control and result in reduced F-actin and increased F-actin at the apical tip of the merozoites respectively. Scale bar represents 1 μm. Treatment of merozoites with actin depolymerizing agents, CytD, ML-B and LA-B, resulted in increased secretion of microneme protein PfAMA-1 (a-c) whereas treatment with polymerized actin stabilizer JAS resulted in reduced secretion of PfAMA-1 (d).Singh S, More KR, Chitnis CE. Role of Calcineurin and Actin Dynamics in Regulated Secretion of Microneme Proteins in Plasmodium falciparum Merozoites during Erythrocyte Invasion. Cell Microbiol. 2013 16(1), 50–63
See original on MMP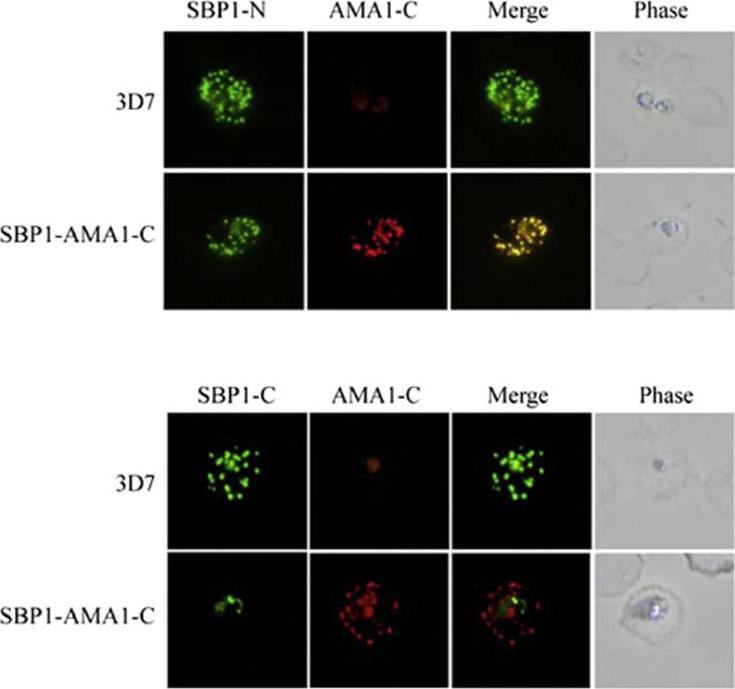
Localisation of SBP1 C-terminal mutants. Immunofluorescence analysis of RBCs infected with either normal (3D7) or transgenic parasites expressing a chimeric SBP1 protein in which the C-terminal domain of SBP1 was replaced with the C-terminal domain of P. falciparum AMA1 (SBP1–AMA1-C). Chimeras localised this protein correctly to Murer’s Clefts. Kats LM, Proellocks NI, Buckingham DW, Blanc L, Hale J, Guo X, Pei X, Herrmann S, Hanssen EG, Coppel RL, Mohandas N, An X, Cooke BM. Interactions between Plasmodium falciparum skeleton-binding protein 1 and the membrane skeleton of malaria-infected red blood cells. Biochim Biophys Acta. 2015 1848(7):1619-1628.
See original on MMP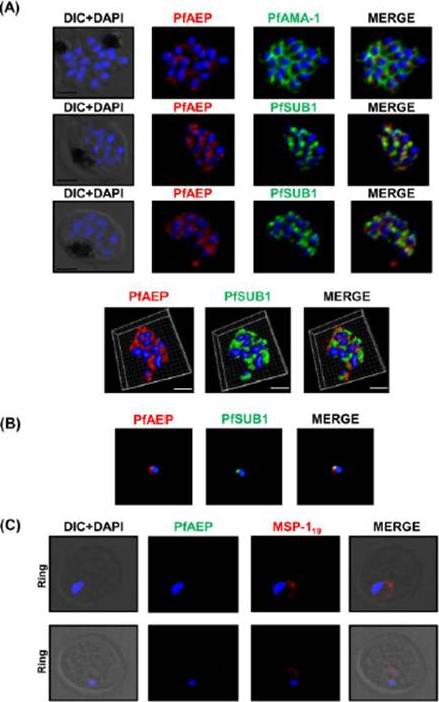
Localization of PfAEP in the exonemes of merozoites (A) Co-localization of PfAEP was studied with micronemal resident protein AMA-1 in bursting schizonts. PfAEP (rabbit serum; red) does not co-localize with AMA-1 (green). The staining for PfAEP co-localized with exonemal marker PfSUB1. Scale bar 2 mm. Three-dimensional reconstruction of the confocal z-stack images of schizonts is shown in panel below Scale bar 1.5mm (B) 2D-structured illumination microscopy (SIM) showed co-localization between the staining of PfAEP and PfSUB1 indicating its localization in the exonemes of P. falciparum merozoites. (C) Young rings were immunostained with anti PfAEP antibody and the antibody against MSP119 as a positive control. No staining was observed for PfAEP in young rings whereas MSP119 showed typical disk like staining around the nucleus.Hans N, Relan U, Dubey N, Gaur D, Chauhan VS. Identification and localization of a Novel Invasin of Plasmodium falciparum. Mol Biochem Parasitol. 2015 Sep 29. [Epub ahead of print]
See original on MMP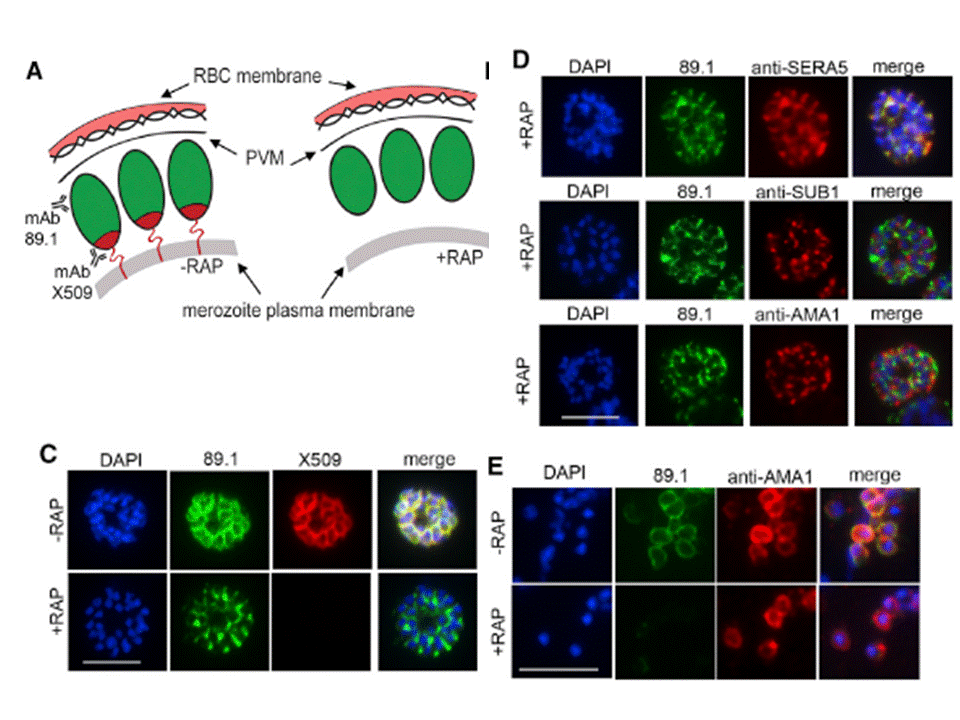
Truncation of MSP1 Produces anEgress Defect (A) Predicted RAP-induced MSP1 truncation in the 3D7MSP1flox42C clones, showing loss of the GPI anchor and C-terminal domain containing the mAb X509 epitope. (C) RAP treatment produces a loss of mAb X509 reactivity and a shift in the IFA pattern of MSP1 to one typical of PV proteins, consistent with the predicted truncation. Numbers of DAPI-stained nuclei did not differ between control and RAPtreated. (D) IFA showing co-localization of truncated MSP1 with SERA5 indicating a PV location. The punctate localization of SUB1 and the microneme protein AMA1 indicates normal organelle biogenesis. (E) IFA showing lack of surface-bound MSP1 on merozoites of RAP-treated 3D7MSP1flox42C1 clone E3. Antibodies to AMA1 (which is expressed on free merozoites) were used as a control.Das S, Hertrich N, Perrin AJ, Withers-Martinez C, Collins CR, Jones ML, Watermeyer JM, Fobes ET, Martin SR, Saibil HR, Wright GJ, Treeck M, Epp C, Blackman MJ. Processing of Plasmodium falciparum Merozoite Surface Protein MSP1 Activates a Spectrin-Binding Function Enabling Parasite Egress from RBCs. Cell Host Microbe. 2015 18(4):433-44.
See original on MMP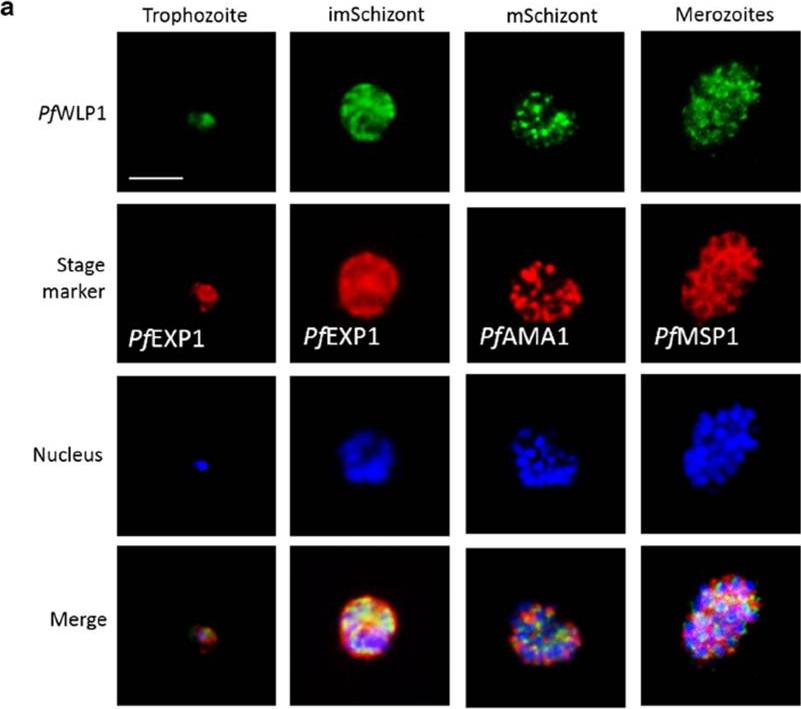
PfWLP1 is expressed in the asexual blood stages of Plasmodium falciparum. a Expression of PfWLP1 in the asexual blood stages. PfWLP1 was immunolabelled with anti-PfWLP1-rp2 antisera (green); the asexual blood stages were visualized with antisera against PfEXP1, PfAMA1 and PfMSP1 (red). The parasite nuclei were highlighted by Hoechst nuclear stain (blue). ImSchizont, immature schizont; mSchizont, mature schizont. Bar 5 μm. Data are representative of five independent experiments each.von Bohl A, Kuehn A, Simon N, Ngongang VN, Spehr M, Baumeister S, Przyborski JM, Fischer R, Pradel G. A WD40-repeat protein unique to malaria parasites associates with adhesion protein complexes and is crucial for blood stageprogeny. Malar J. 2015 Nov 4;14(1):435.
See original on MMP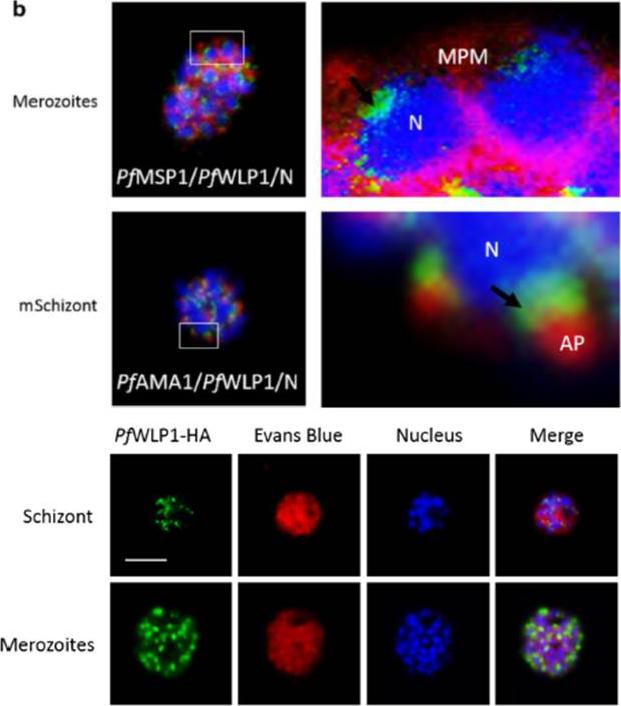
PfWLP1 is expressed in the asexual blood stages of Plasmodium falciparum. Upper panel: Localization of PfWLP1 underneath the apical pole of merozoites. Image enlargements (right) of mature schizonts (left), labelled with antibodies against PfWLP1 (green) and PfMSP1 or PfAMA1 (red) and stained with Hoechst (blue). AP apical pole, N nucleus, MPM merozoite plasma membrane. Bar 5 μm. Data are representative of five independent experiments each. Lower panel: Expression of PfWLP1-HA in asexual blood stages. PfWLP1 expression in the PfWLP1-HA asexual blood stages. PfWLP1-HA was immunolabelled with anti-HA antisera (green); the asexual blood stages were visualized by Evans Blue counterstaining (red). The parasite nuclei were highlighted by Hoechst nuclear stain (blue),von Bohl A, Kuehn A, Simon N, Ngongang VN, Spehr M, Baumeister S, Przyborski JM, Fischer R, Pradel G. A WD40-repeat protein unique to malaria parasites associates with adhesion protein complexes and is crucial for blood stage progeny. Malar J. 2015 Nov 4;14(1):435.
See original on MMP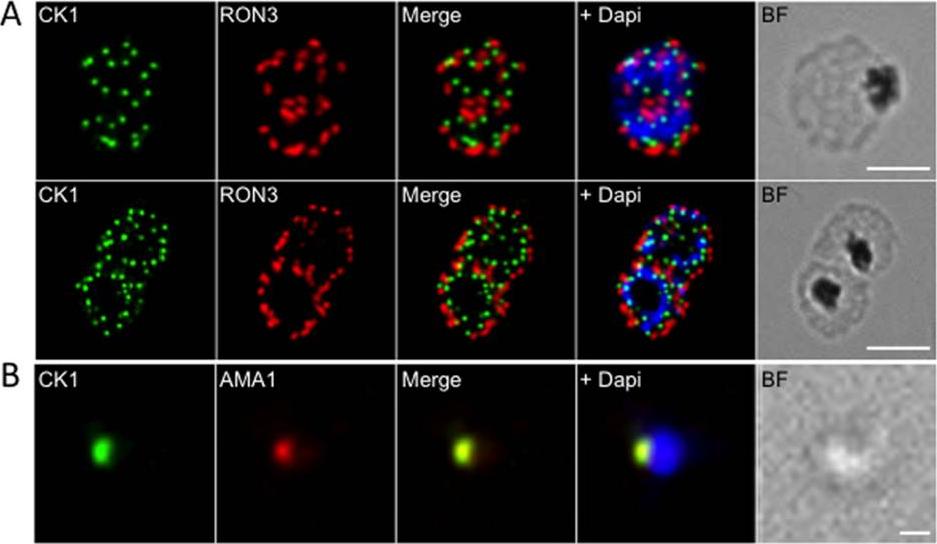
Localisation of PfCK1 in micronemes. (A) Localisation of PfCK1 and PfRON3 in schizonts and segmenters. DAPI was used to stain the nucleus and the scale bar represents 10μm. (B) Localisation of PfCK1 and AMA-1 in merozoites. DAPI was used to stain the nucleus and the scale bar represents 1μm. A punctate pattern emerges in segmenters, with one PfCK1 dot in each developing merozoite, adjacent to the rhoptry marker RON3. In line with this observation, PfCK1 colocalises with AMA-1, a protein of micronemes that is released at the very early stages of invasion.Dorin-Semblat D, Demarta-Gatsi C, Hamelin R, Armand F, Carvalho TG, Moniatte M, Doerig C. Malaria Parasite-Infected Erythrocytes Secrete PfCK1, the Plasmodium Homologue of the Pleiotropic Protein Kinase Casein Kinase 1. PLoS One. 2015 10(12):e0139591.
See original on MMPMTRAP is cleaved within the micronemes. A) IFA of compound 1 (C1) treated schizonts co-labelled with mAMA1 antibodies (green) and rM-tsr (top panels), rM-mid (middle panels) or rM-tail (bottom panels) (red in respective images). Scale bars = 1 μm. C) Schematic of pre- (left half) and post- (right half) invasion merozoites, showing the predominant distribution of each protein tested as part of this study. It should be noted that all proteins show a degree of variability in labelling distributions across merozoites imaged to an extent that is not possible to depict here.Riglar DT, Whitehead L, Cowman AF, Rogers KL, Baum J. Localization-based imaging of malarial antigens during red cell entry reaffirms role for AMA1 but not MTRAP in invasion. J Cell Sci. 2015. [Epub ahead of print]
See original on MMP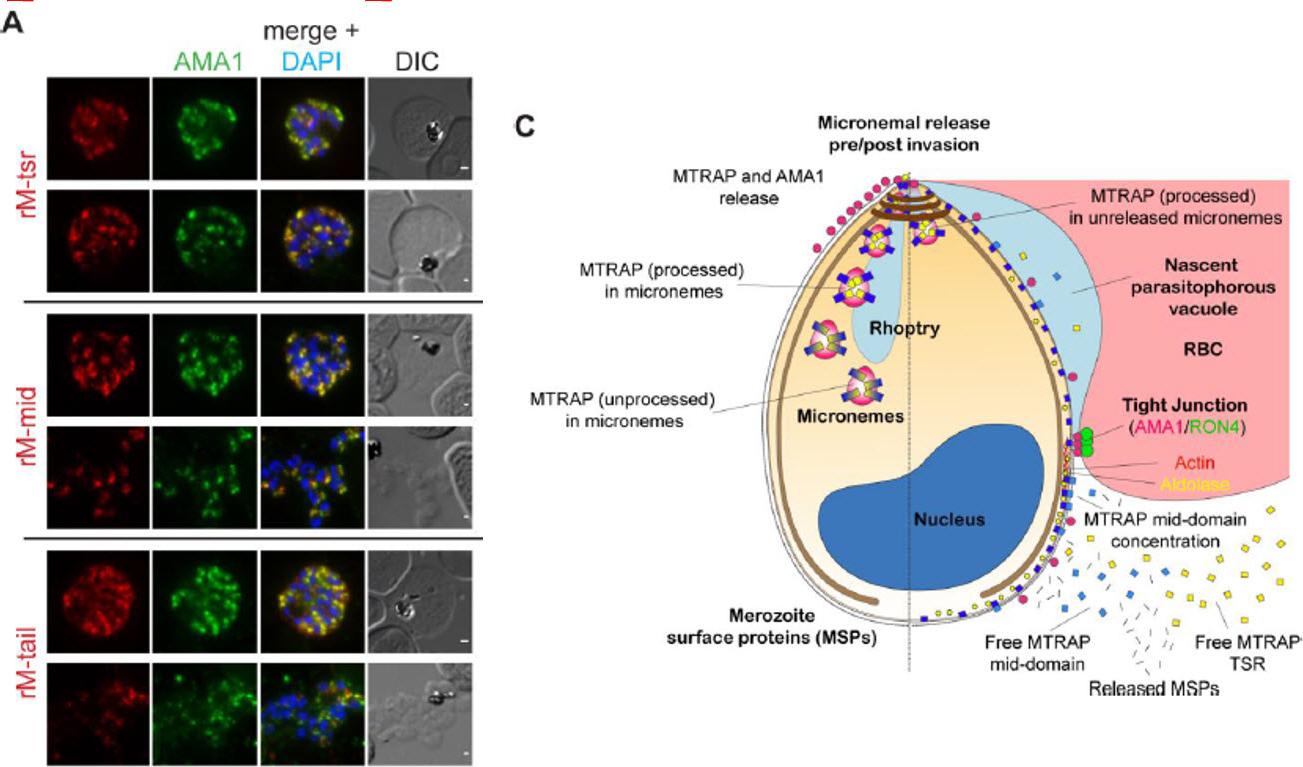
MTRAP is cleaved within the micronemes. A) IFA of compound 1 (C1) treated schizonts co-labelled with mAMA1 antibodies (green) and rM-tsr (top panels), rM-mid (middle panels) or rM-tail (bottom panels) (red in respective images). Scale bars = 1 μm. C) Schematic of pre- (left half) and post- (right half) invasion merozoites, showing the predominant distribution of each protein tested as part of this study. It should be noted that all proteins show a degree of variability in labelling distributions across merozoites imaged to an extent that is not possible to depict here.Riglar DT, Whitehead L, Cowman AF, Rogers KL, Baum J. Localization-based imaging of malarial antigens during red cell entry reaffirms role for AMA1 but not MTRAP in invasion. J Cell Sci. 2015. [Epub ahead of print]
See original on MMP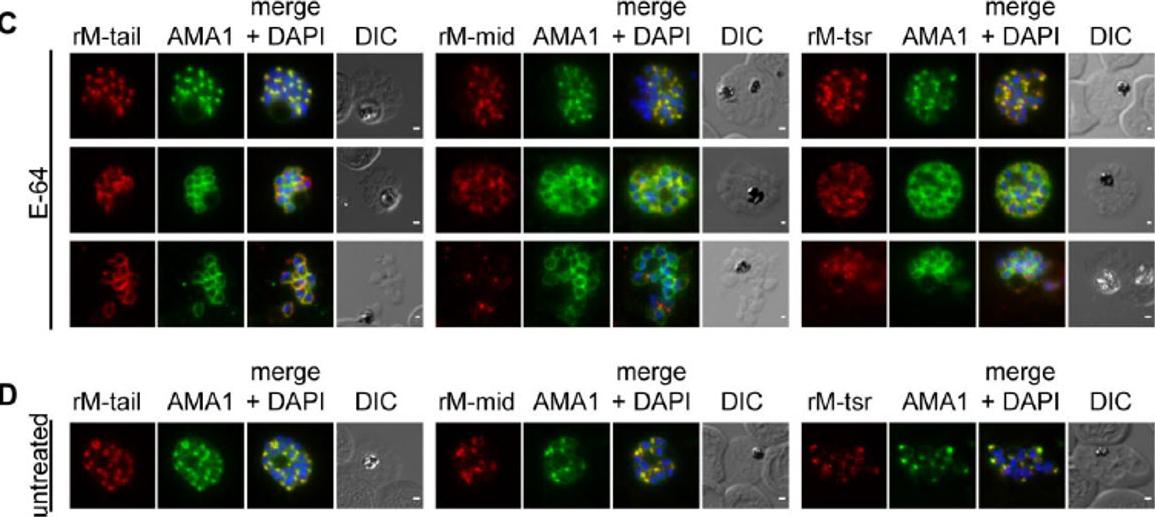
MTRAP is cleaved irrespective of both invasion and egress from the schizont. (C) treated with E-64 to prevent egress or (D) from untreated culture, co-labelled AMA1 monoclonal antibodies (green) and rM-tail (left), rM-mid (middle) or rM-tsr (right) (red in respective images). Scale bars = 1 μm.Riglar DT, Whitehead L, Cowman AF, Rogers KL, Baum J. Localization-based imaging of malarial antigens during red cell entry reaffirms role for AMA1 but not MTRAP in invasion. J Cell Sci. 2015. [Epub ahead of print]
See original on MMP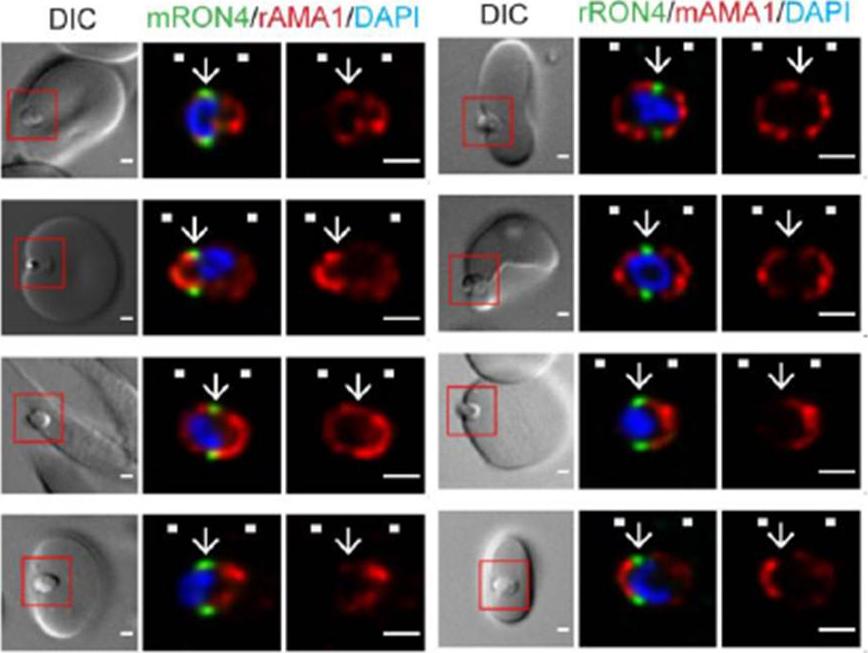
Longitudinal intensity profiling demonstrates antibody specific variation in AMA1 localisation at the tight junction. Single-slice images of example merozoites (left panel) mRON4 (green) vs rAMA1 (red) or(right panel) rRON4 (green) vs mAMA1 (red). Scale bars = 1 μm. Red boxes in DIC images indicate zoomed regions for middle and right panels. Arrows indicate the position of the RON4 labelled tight junctionRiglar DT, Whitehead L, Cowman AF, Rogers KL, Baum J. Localization-based imaging of malarial antigens during red cell entry reaffirms role for AMA1 but not MTRAP in invasion. J Cell Sci. 2015. [Epub ahead of print]
See original on MMP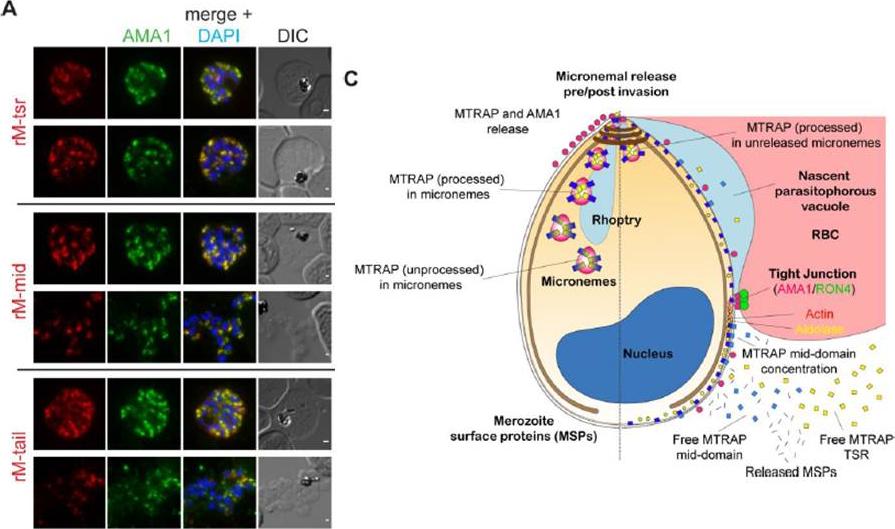
MTRAP is cleaved within the micronemes. A) IFA of compound 1 (C1) treated schizonts co-labelled with mAMA1 antibodies (green) and rM-tsr (top panels), rM-mid (middle panels) or rM-tail (bottom panels) (red in respective images). Scale bars = 1 μm. C) Schematic of pre- (left half) and post- (right half) invasion merozoites, showing the predominant distribution of each protein tested as part of this study. It should be noted that all proteins show a degree of variability in labelling distributions across merozoites imaged to an extent that is not possible to depict here.Riglar DT, Whitehead L, Cowman AF, Rogers KL, Baum J. Localization-based imaging of malarial antigens during red cell entry reaffirms role for AMA1 but not MTRAP in invasion. J Cell Sci. 2015. [Epub ahead of print]
See original on MMP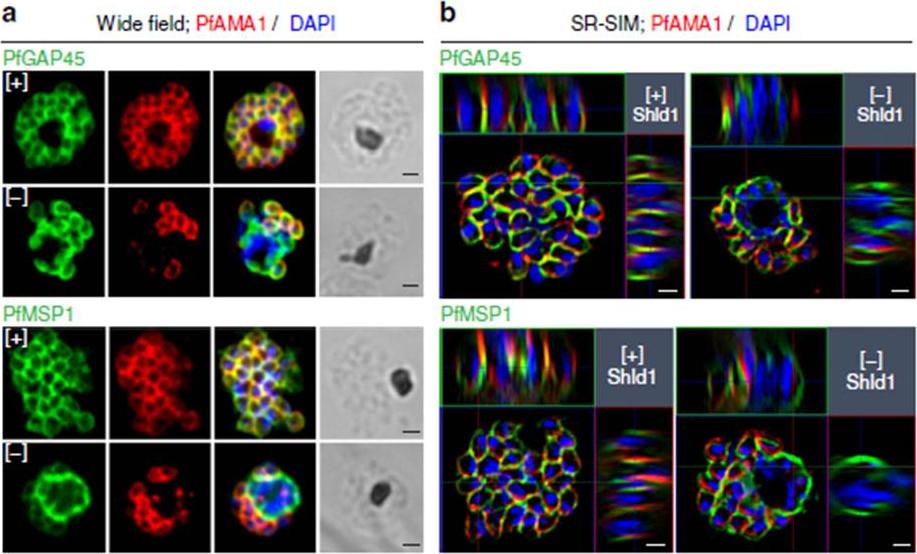
PfMOP deficiency leads to incomplete formation of the IMC. (a) Wide-field and (b) SR-SIM (super-resolution structured illumination microscopy) of representative pictures of E64-treated [+]/[-] Shld1 schizont stage PfMOP-DD parasites using antibodies against PfGAP45 and PfMSP1. Scale bar, 1 mm.Absalon S, Robbins JA, Dvorin JD. An essential malaria protein defines the architecture of blood-stage and transmission-stage parasites. Nat Commun. 2016 7:11449.
See original on MMP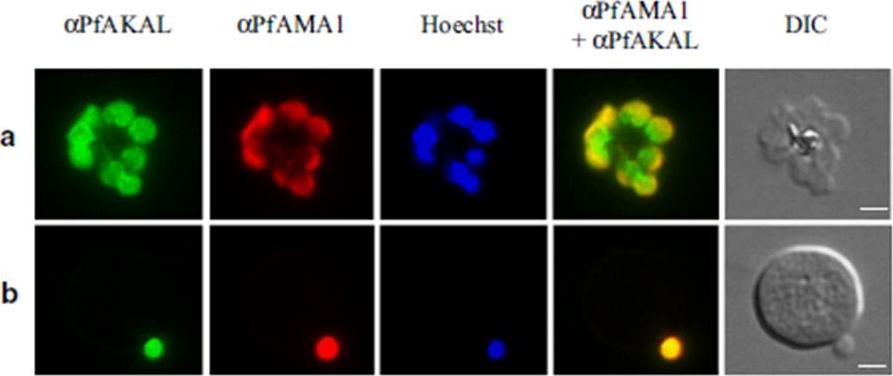
PfAKAL localization in P. falciparum merozoites by indirect immunofluorescence assays. Images with PfAKAL localization (green), PfAMA1 localization (red), Hoechst DNA staining (blue) and differential interference contrast (DIC) and were taken. a P. falciparum 3D7 egressing merozoites, b P. falciparum 3D7 invading merozoite. The bars represent 2 μm. PfAKAL is expressed in egressing (a) and invading (b) merozoites.Bandje K, Naissant B, Bigey P, Lohezic M, Vayssières M, Blaud M, Kermasson L, Lopez-Rubio JJ, Langsley G, Lavazec C, Deloron P, Merckx A. Characterization of an A-kinase anchoring protein-like suggests an alternative way of PKA anchoring in Plasmodium falciparum. Malar J. 2016 15(1):248.
See original on MMP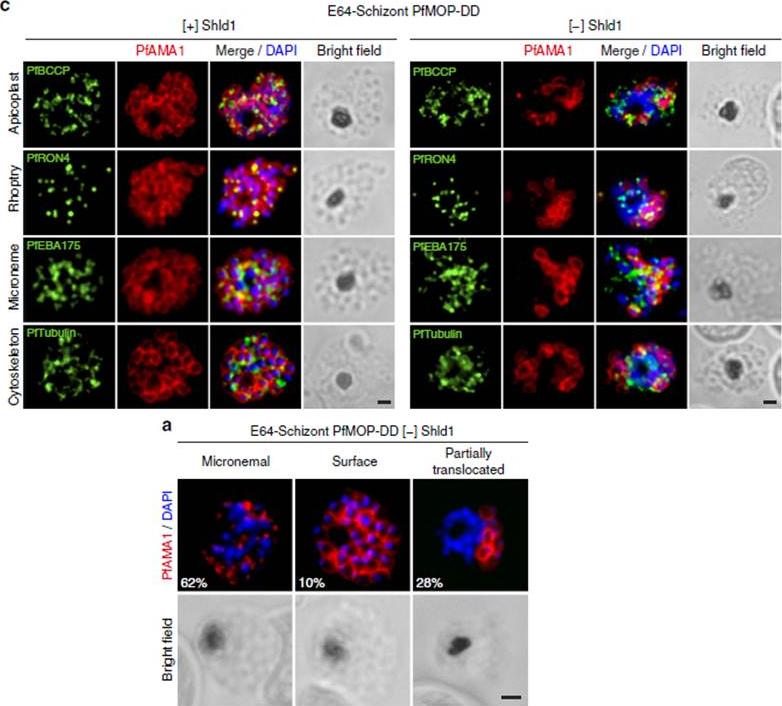
PfAMA1 translocation is aberrant in schizonts with PfMOP-knockdown. (a) Schizonts from [+]/[-] Shld1 PfMOP-DD parasites were E64-treated, fixed, probed with anti-PfAMA1 and scored as micronemal (M), partially translocated (PT) or surface (S), representative [+] Shld1 parasite IFAs shown. (c) Synchronized schizont stage (40–44 h) parasites, maintained with 250nM (left panel) or 0 nM (right panel) Shld1, were incubated 6 h in presence of 10 mM E64, methanol-fixed, permeabilized, and stained using antibodies against PfRON4, PfRhopH3, PfEBA175 and PfTubulin. Staining for these markers was similar in [+] and [-] Shld1 conditions. PfAMA1 staining was used to identify E64-treated schizonts that were sufficiently mature (that is, surface staining or partially translocated staining, but not micronemal staining, scale bar, 1 mm).Absalon S, Robbins JA, Dvorin JD. An essential malaria protein defines the architecture of blood-stage and transmission-stage parasites. Nat Commun. 2016 7:11449.
See original on MMP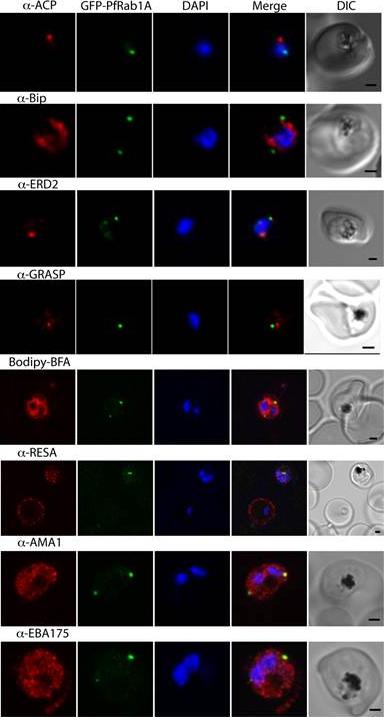
GFP-PfRab1A fluorescence does not co-localize with ER/Golgi, dense granules or microneme markers. GFP-PfRab1A fluorescence is distinct from the localization of markers for the apicoplast (ACP), the ER (Bip), the Golgi (ERD2 and GRASP), as well as from staining of the ER/Golgi with Bodipy BFA. GFP-Rab1A fluorescence is also distinct from the localization of markers for dense granules (RESA) or micronemes (AMA1 and EBA175).Morse D, Webster W, Kalanon M, Langsley G, McFadden GI. Plasmodium falciparum Rab1A Localizes to Rhoptries in Schizonts. PLoS One. 2016 Jun 27;11(6):e0158174
See original on MMP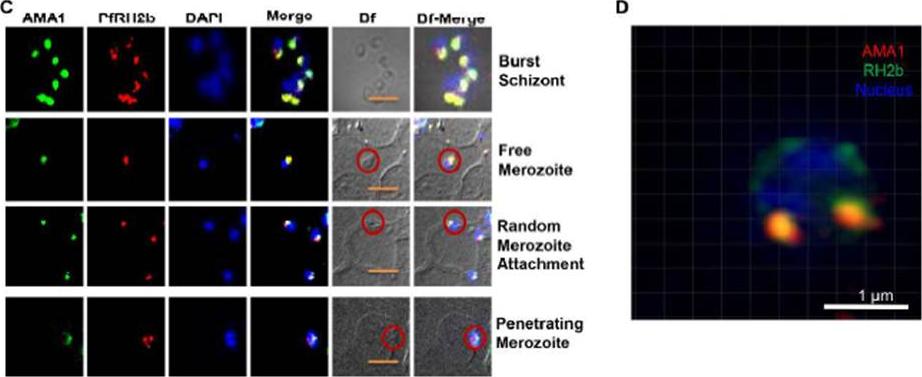
PfRH2b is associated with the moving junction during merozoite invasion. The process of merozoite invasion was closely monitored to locate the position of PfRH2b of burst schizont, free merozoites, random merozoite attachment, apposed merozoite and penetrating merozoite. The presence of PfRH2b (PfRH2b, red) at the moving junction was further confirmed with AMA1 (C, green) as common markers with punctuate staining. Merozoites of focus are shown with red circles. The bright field images labeled as Bf. Scale bar indicates 2 μm. (D) A sliceof a three-dimensional structured illumination microscopy (3D-SIM) on merozoite at the junction further confirmed the punctuated pattern of PfRH2b (green) to be in close proximity to AMA1 (red). Scale bar indicates 1 μm. In the merged images, areas of overlap between the red and the green signals are shown inyellow. DAPI (blue) stained for nuclei.Aniweh Y, Gao X, Gunalan K, Preiser PR. PfRH2b Specific Monoclonal Antibodies Inhibit Merozoite Invasion. Mol Microbiol. 2016 Jul 20.
See original on MMP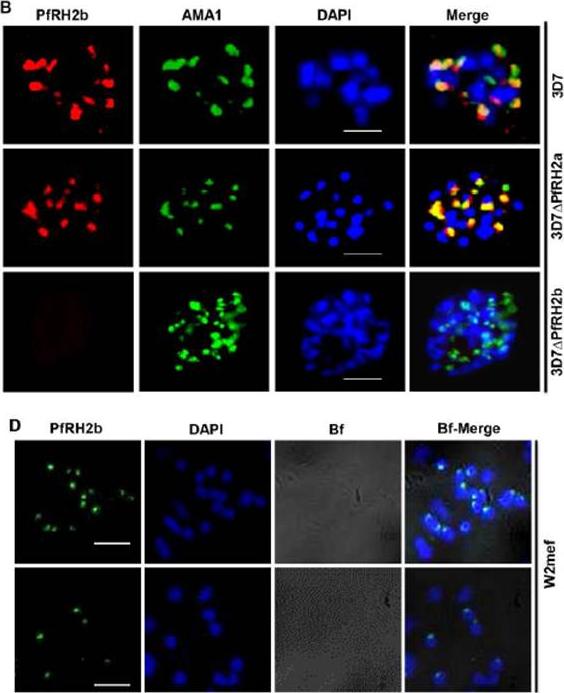
(B) IFA indicates the specificity of mAb A7 in schizont stage parasites. PfRH2b (PfRH2b, red) co-stained with AMA1 (AMA1, green), DAPI for nuclei (blue) in 3D7, 3D7ΔPfRH2a and 3D7ΔPfRH2b schizont. In the merged images, areas of overlap between the red and the green signals are shown in yellow. Scale bar indicated in white represents 2 μm. (D) Nonpermeabilized W2mef merozoites show surface expression of PfRH2b using mAb A7. PfRH2b shown green (RH2b A7), DAPI for nuclei (blue) and bright field (Bf) images shown in gray. Scale bar indicates 2 μm.Aniweh Y, Gao X, Gunalan K, Preiser PR. PfRH2b Specific Monoclonal Antibodies Inhibit Merozoite Invasion. Mol Microbiol. 2016 Jul 20.
See original on MMP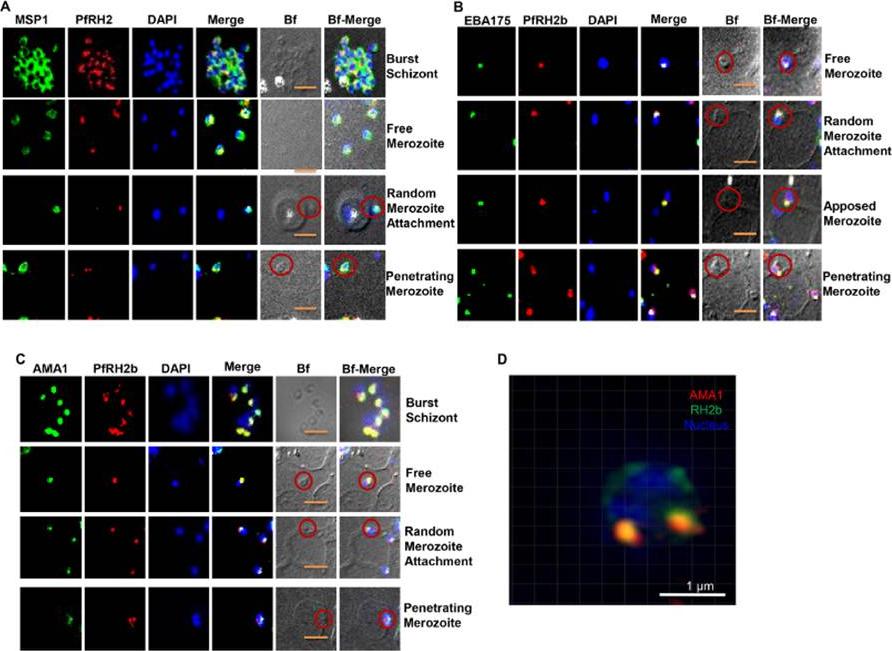
PfRH2b is associated with the moving junction during merozoite invasion. The process of merozoite invasion was closely monitored to locate the position of PfRH2b of burst schizont, free merozoites, random merozoite attachment, apposed merozoite and penetrating merozoite. A. Merozoites were stained for MSP1 (MSP1, green) and PfRH2 (PfRH2, red). PfRH2 apical punctuate staining was observed in the free and attaching merozoites while in invading merozoites two punctuate staining were observed at the anchorage of the moving junction. The presence of PfRH2b (PfRH2b, red) at the moving junction was further confirmed with EBA175 (B, green) and AMA1 (C, green) as common markers with punctuate staining. Merozoites of focus are shown with red circles. The bright field images labelled as Bf. Scale bar indicates 2 lm. D. A slice of a three-dimensional structured illumination microscopy (3D-SIM) on merozoite at the junction further confirmed the punctuated pattern of PfRH2b (green) to be in close proximity to AMA1 (red). Scale bar indicates 1 lm. In the merged images, areas of overlap between the red and the green signals are shown in yellow. DAPI (blue) stained for nuclei.Aniweh Y, Gao X, Gunalan K, Preiser PR. PfRH2b specific monoclonal antibodies inhibit merozoite invasion. Mol Microbiol. 2016 Nov;102(3):386-404.
See original on MMP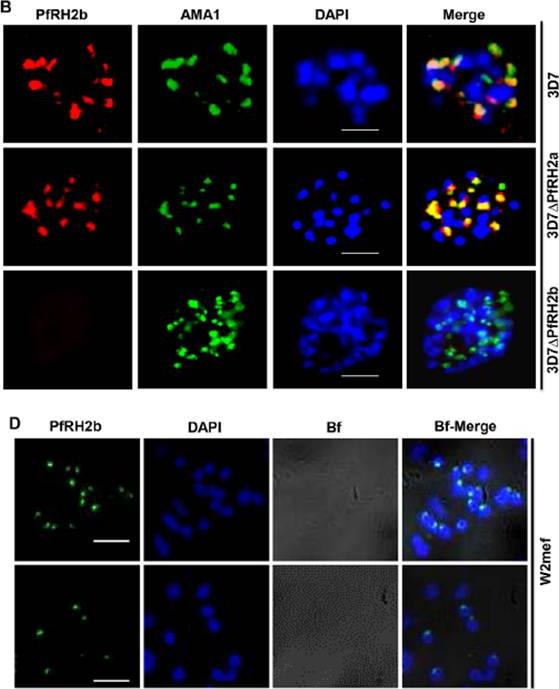
B. IFA indicates the specificity of mAb A7 in schizont stage parasites. PfRH2b (PfRH2b, red) co-stained with AMA1 (AMA1, green), DAPI for nuclei (blue) in 3D7, 3D7DPfRH2a and 3D7DPfRH2b schizont. In the merged images, areas of overlap between the red and the green signals are shown in yellow. Scale bar indicated in white represents 2 mm. D. Non-permeabilized W2mef merozoites show surface expression of PfRH2b using mAb A7. PfRH2b shown green (RH2b A7), DAPI for nuclei (blue) and bright field (Bf) images shown in gray. Scale bar indicates 2 mm. (3D7DPfRH2a (clone in which PfRH2a has been deleted), 3D7DPfRH2b (clone in which the PfRH2b gene has been deleted) parasites. The punctate staining pattern observed for A7 was in close proximity to the staining observed with an antibody targeting the micronemal protein Apical membrane antigen 1 (AMA1), indicating an apical location of PfRH2b within the merozoite.Aniweh Y, Gao X, Gunalan K, Preiser PR. PfRH2b specific monoclonal antibodies inhibit merozoite invasion. Mol Microbiol. 2016 Nov;102(3):386-404.
See original on MMP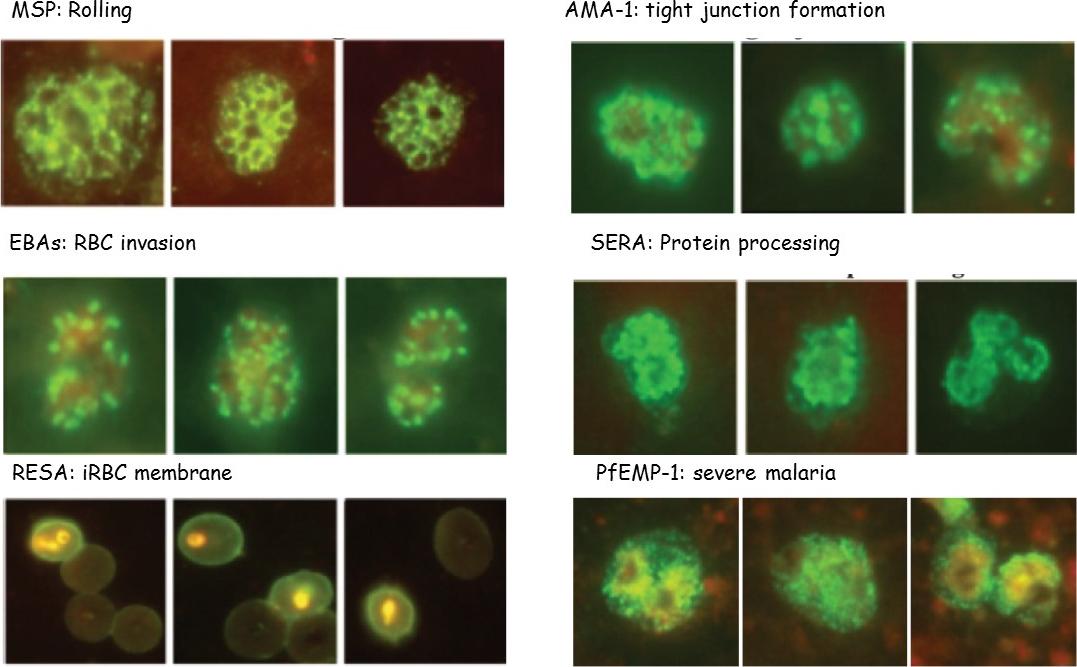
P. falciparum merozoite protein location is shown, as determined by IFA and functions: rolling on the membrane by MSPs, TJ formation mediated by AMA-1-and apical rhoptries proteins, RBC deformation and invasion by microneme EBAs, protein processing by intracytoplasmic SERA-5, iRBC membrane expression by RESA and severe malaria (SM) and echinocytosis by PfEMP-1.Patarroyo ME, Arevalo-Pinzon G, Reyes C, Moreno-Vranich A, Patarroyo MA. Malaria Parasite Survival Depends on Conserved Binding Peptides' Critical Biological Functions. Curr Issues Mol Biol. 2016;18:57-78.
See original on MMP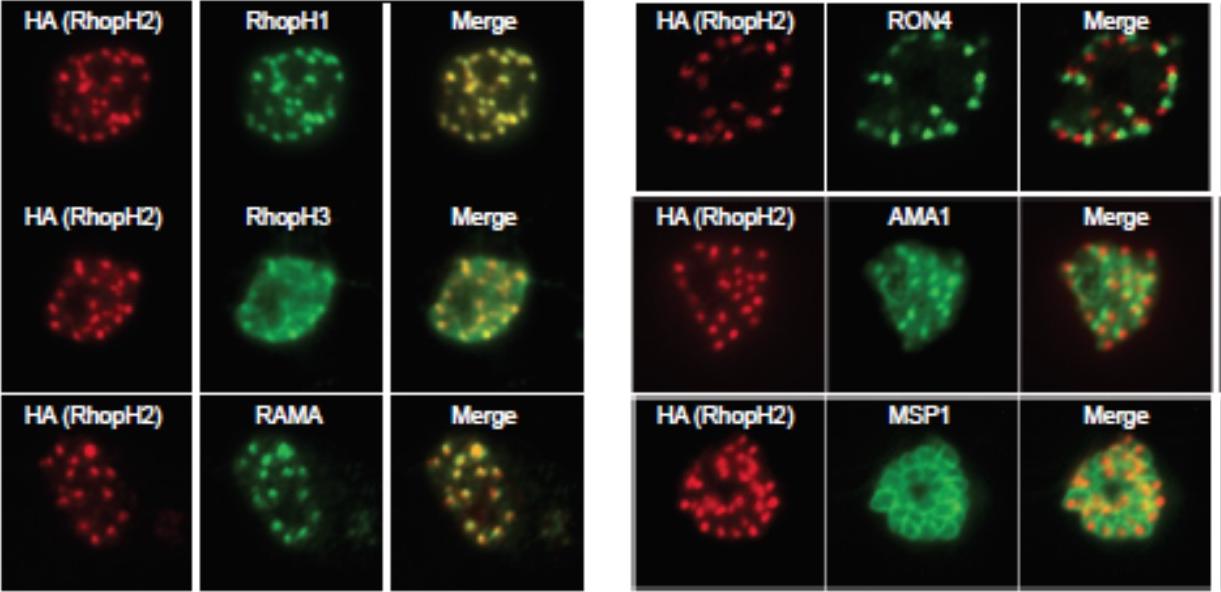
Generation of transgenic parasites in which RhopH2 is epitope-tagged. Immunofluorescence analysis (IFA) on schizonts fixed with acetone/methanol and labelled with anti-HA antibody to detect RhopH2 and other antibodies, as indicated. Immunofluorescence analysis (IFA) confirmed RhopH2-HA localized to the rhoptry and co-localized with other rhoptry bulb proteins, RhopH1, RhopH3 and RAMA but not with the rhoptry neck protein, RON4, the micronemal marker, AMA-1 or the plasma membrane protein MSP1.Counihan N, Chisholm SA, Bullen HE, Srivastava A, Sanders PR, Jonsdottir TK, Weiss GE, Ghosh S, Crabb BS, Creek DJ, Gilson PR, de Koning-Ward TF. Plasmodium falciparum parasites deploy RhopH2 into the host erythrocyte to obtain nutrients, grow and replicate. Elife. 2017 Mar 2;6. pii: e23217
See original on MMP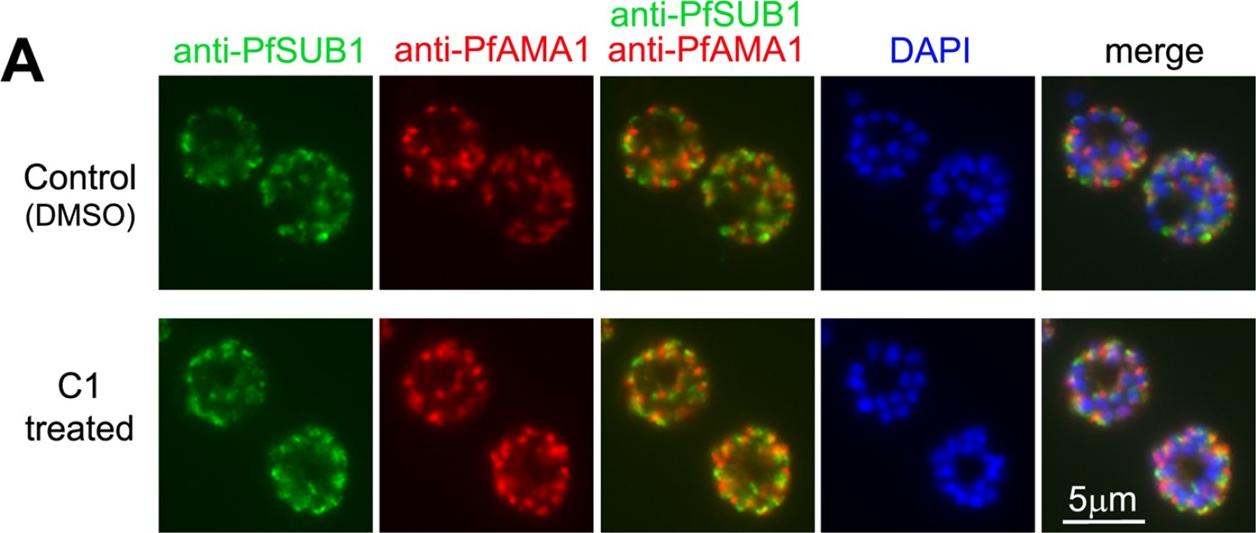
C1 and C2 do not affect expression, trafficking or maturation of PfSUB1. (A) IFA of mature wt 3D7 parasites following treatment for 6 h with DMSO only (1% v/v) or 2.5 mM C1, probed with the anti-PfSUB1 mAb NIMP.M7 or a rabbit anti-PfAMA1 serum. The PfPKG inhibitor had no effect on localisation or expression of PfSUB1 or PfAMA1. For clarity, the anti-PfSUB1 and anti-PfAMA1 signals are shown merged in addition to a final merge with the DAPI signal. Similar results were obtained following treatment with 1.5 mM C2 (not shown). Note that treatment with the PKG inhibitors also had no effect on schizont replication as determined by counts of the average number of nuclei per schizont (data not shown, but compare DAPI staining of untreated and treated schizonts). Collins CR, Hackett F, Strath M, Penzo M, Withers-Martinez C, Baker DA, Blackman MJ. Malaria parasite cGMP-dependent protein kinase regulates blood stage merozoite secretory organelle discharge and egress. PLoS Pathog. 2013 9(5):e1003344.
See original on MMP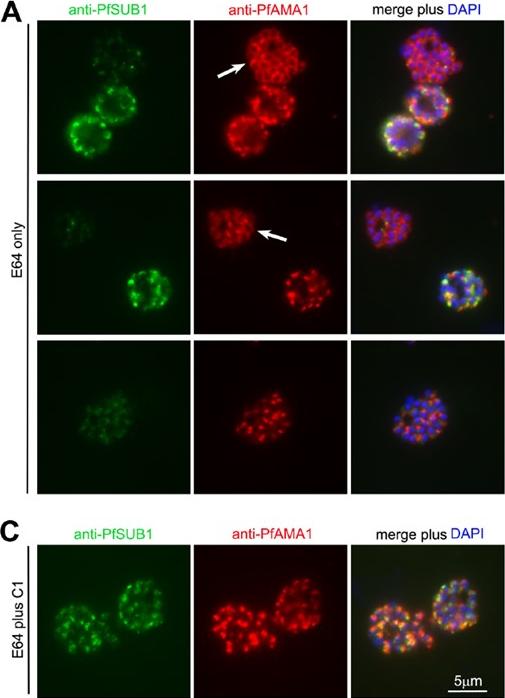
PfPKG inhibitors block discharge of exonemes and micronemes. (A) IFA of wt 3D7 parasites allowed to develop beyond the point of egress in the presence of E64 only (50 mM). Top two rows: segmented schizonts displaying a merozoite surface localisation of PfAMA1 (arrowed; ,45% of the schizont population) as a result of its discharge from micronemes, exhibit a weak PfSUB1 signal compared to less mature schizonts in which both the PfAMA1 and PfSUB1 signals remain punctate. The bottom row of images in (A) shows a schizont in which the PfAMA1 signal is intermediate between punctate and merozoite surface. (C) IFA of wt 3D7 parasites allowed to develop beyond the point of egress in the presence of both E64 and C1. No discharge or relocalisation of PfSUB1 or PfAMA1 was evident in counts of .5,000 schizonts from a total of 3 independent experiments. Identical results were obtained with E64 plus C2 (not shown).Collins CR, Hackett F, Strath M, Penzo M, Withers-Martinez C, Baker DA, Blackman MJ. Malaria parasite cGMP-dependent protein kinase regulates blood stage merozoite secretory organelle discharge and egress. PLoS Pathog. 2013 9(5):e1003344.
See original on MMP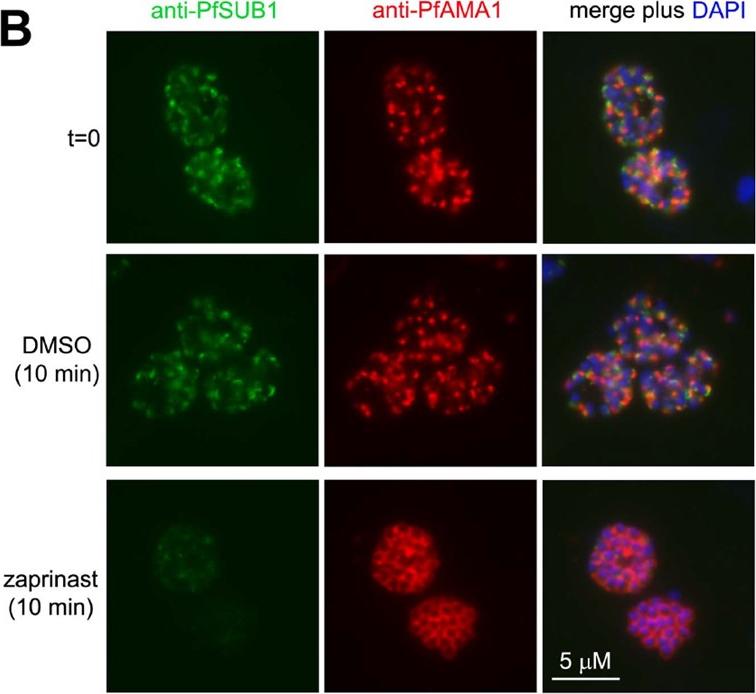
Zaprinast (PKG inhibitor) induces rapid discharge of exonemes and micronemes. (B) Zaprinast treatment of schizonts (in the presence of E64) induces rapid relocalisation of PfAMA1 and loss of PfSUB1. Fixed, permeabilized schizonts were examined by IFA with the antibodies indicated. Zaprinast-induced relocalisation of PfSUB1 and PfAMA1 was evident even in immature (non-segmented) schizonts, but here took the form of translocation of both proteins to a striking circumferential punctate pattern, perhaps indicating accumulation of immature micronemes and exonemes at the periphery of the schizont.Collins CR, Hackett F, Strath M, Penzo M, Withers-Martinez C, Baker DA, Blackman MJ. Malaria parasite cGMP-dependent protein kinase regulates blood stage merozoite secretory organelle discharge and egress. PLoS Pathog. 2013 9(5):e1003344.
See original on MMP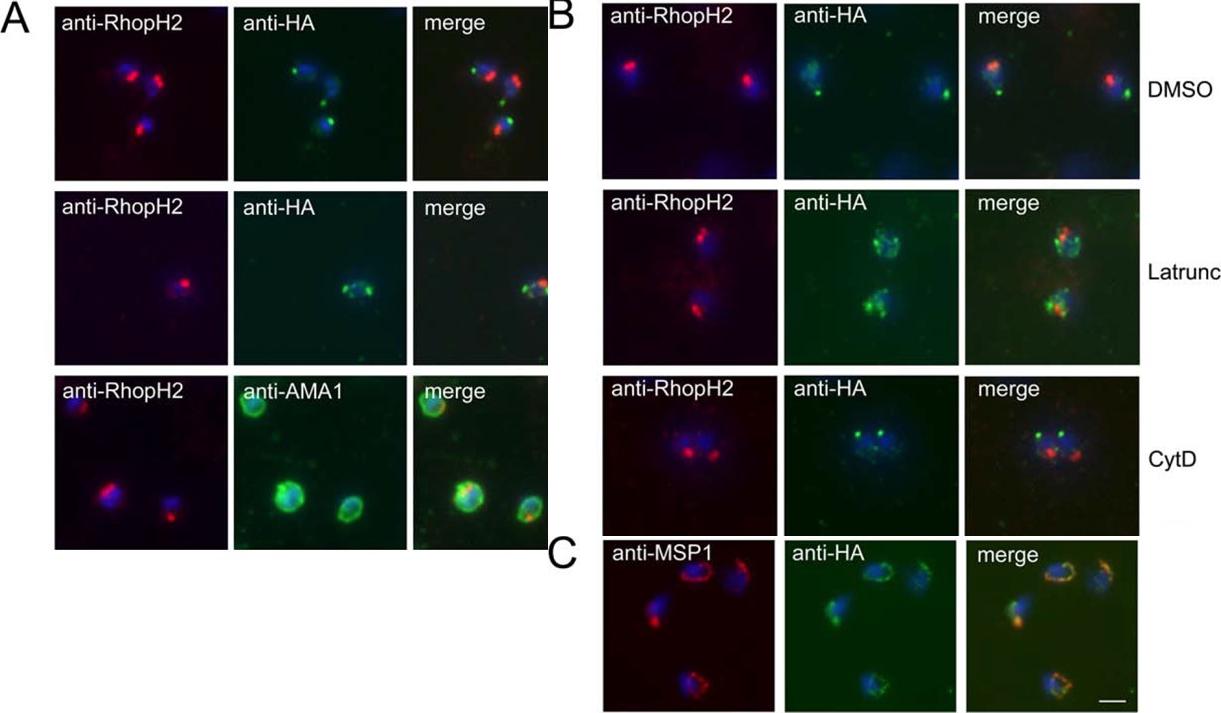
PfSUB2 Redistributes to the Free Merozoite Posterior in an Actin-Dependent Manner. (A) IFA images of free merozoites of PfSUB2HA clone 2D dual-labelled with mAb 61.3 (anti-RhopH2) and either mAb 3F10 (anti-HA) or 4G2 (anti-AMA1). The anti-HA signal formed either a single punctate signal at the extreme posterior of the merozoite (top), or twin foci lateral to its anterior–posterior axis (middle). Identical results were obtained using PfSUB2HA clone 10E, or using anti-RAP2 mAb H5 as the rhoptry marker (not shown). (B) Latrunculin inhibits rearward translocation of PfSUB2. Merozoites of PfSUB2HA clone 2D released into medium containing 1% (v/v) DMSO only (solvent control), or 5 mM latrunculin A (Latrunc), or 4 mM cytochalasin D (Cyt D), probed as above with mAb 61.3 (anti-RhopH2) and mAb 3F10 (anti-HA). Note that neither compound had any effect on the efficiency of schizont rupture and merozoite release, nor on AMA1 relocalisation, but both blocked erythrocyte invasion by more than 95%at the concentrations used (not shown). (C) PfSUB2 remains at the plasma membrane of the newly invaded parasite. Ring stages of PfSUB2HA clone 2D (<2 h post-invasion) probed with anti-MSP1 mAb 1E1 and anti-HA mAb 3F10. Parasite nuclei are stained throughout with DAPI (blue). Scale bar represents 2 mm.Harris PK, Yeoh S, Dluzewski AR, O'Donnell RA, Withers-Martinez C, Hackett F, Bannister LH, Mitchell GH, Blackman MJ. Molecular identification of a malaria merozoite surface sheddase. PLoS Pathog. 2005 Nov;1(3):241-51.
See original on MMP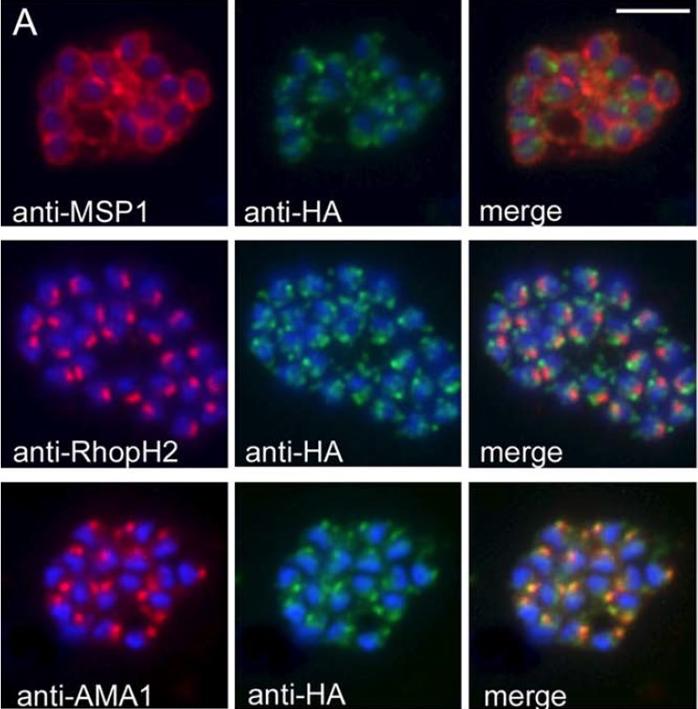
PfSUB2 Is a Microneme Protein. (A) IFA images of schizonts of PfSUB2HA clone 2D dual-labelled with mAbs X509 (anti-MSP1), 61.3 (anti-RhopH2), or 4G2 (anti-AMA1), plus in each case mAb 3F10 (anti-HA) The anti-HA signal co-localised only with the anti-AMA1 signal. Identical results were obtained with the unclonedtransgenic PfSUB2HA line, and/or when anti-RAP2 mAb H5 was used as the rhoptry marker instead of mAb 61.3 (not shown). Parasite nuclei are stained throughout with DAPI (blue). Scale bar represents 2 mm. the anti-HA signal was quite different from that of MSP1, it was adjacent to but distinct from that of the rhoptry specific signals, but it co-localised completely with that ofAMA1, indicating that in mature schizonts, PfSUB2 accumulates in micronemes.Harris PK, Yeoh S, Dluzewski AR, O'Donnell RA, Withers-Martinez C, Hackett F, Bannister LH, Mitchell GH, Blackman MJ. Molecular identification of a malaria merozoite surface sheddase. PLoS Pathog. 2005 Nov;1(3):241-51.
See original on MMP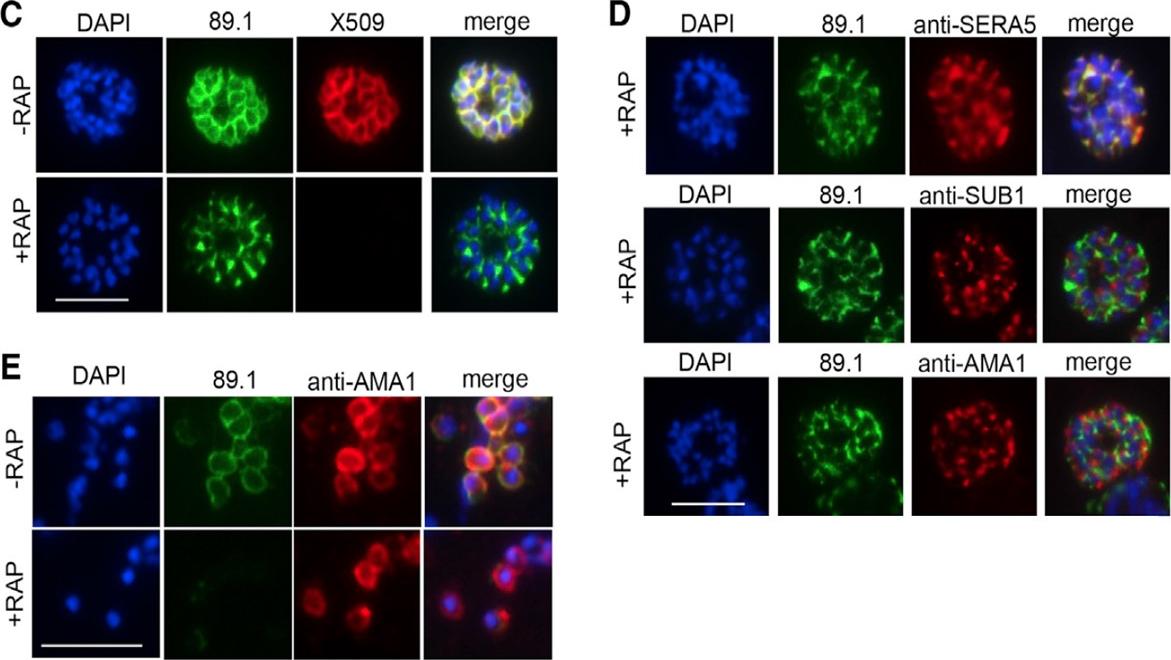
Truncation of MSP1 Produces an Egress Defect. (C) RAP treatment produces a loss of mAb X509 reactivity and a shift in the IFA pattern of MSP1 to one typical of PV proteins, consistent with the predicted truncation. Numbers of DAPI-stained nuclei did not differ between control and RAP treated schizonts (mean values: 21.2 ± 3.4 and 20.6 ± 4.0 nuclei per schizont, respectively, n = 24). (D) IFA showing co-localization of truncated MSP1 with SERA5 indicating a PV location. The punctate localization of SUB1 and the microneme protein AMA1 indicates normal organelle biogenesis. (E) IFA showing lack of surface-bound MSP1 on merozoites of RAP-treated 3D7MSP1flox42C1 clone E3. Antibodies to AMA1 (which is expressed on free merozoites) were used as a control. Analysis of RAP-treated 3D7MSP1flox42C parasites showed highly efficient excision, resulting in exclusive expression of truncated MSP1 in mature schizonts at the end of the same erythrocytic cycle (C). No effects on merozoite development were discernible. The modified MSP1 was trafficked to the PV as expected for a non-membrane bound merozoite surface protein (C–D) but was not present on the surface of free merozoites (E)Das S, Hertrich N, Perrin AJ, Withers-Martinez C, Collins CR, Jones ML, Watermeyer JM, Fobes ET, Martin SR, Saibil HR, Wright GJ, Treeck M, Epp C, Blackman MJ. Processing of Plasmodium falciparum Merozoite Surface Protein MSP1 Activates a Spectrin-Binding Function Enabling Parasite Egress from RBCs. Cell Host Microbe. 2015 18(4):433-44. PMID: 26468747
See original on MMP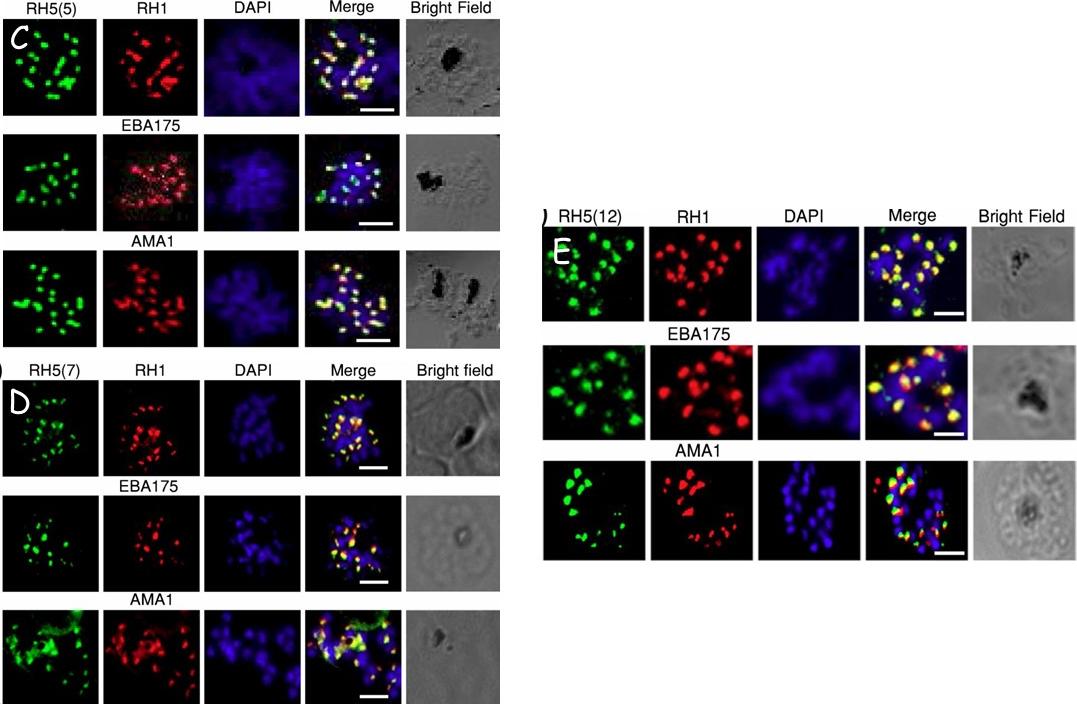
Validation of RH5 mAbs by Immunofluroscence Microscopy. Smears of 3D7 late stage schizont were co- stained with RH5(5) (c) (7) (d) and (12) (e) (first column, green) and antibodies against other invasion markers RH1, EBA175 and AMA1 respectively (second column, red). Parasite nuclei were stained with DAPI (blue). In the merged images, areas of overlap between the red and the green signals are shown in yellow. Bright field indicates the outline of the schizont. Scale bar =2μm.Aniweh Y, Gao X, Hao P, Meng W, Lai SK, Gunalan K, Chu TT, Sinha A, Lescar J, Chandramohanadas R, Li HY, Sze SK, Preiser PR. RH5-Basigin interaction induceschanges in the cytoskeleton of the host RBC. Cell Microbiol. 2017 Apr 13 [Epub ahead of print] PMID: 28409866
See original on MMP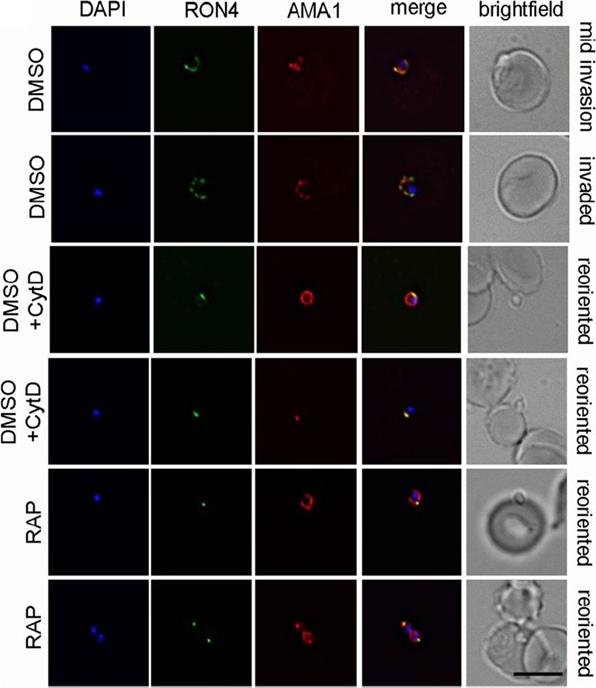
PfACT1 KO merozoites can secrete their micronemes and form a tight junction (TJ) but cannot invade erythrocytes. IFA of TJ formation. Colocalisationof rhoptry neck protein 4 (RON4) and AMA1 at the merozoite-erythrocyte boundary indicates successful TJ formation in DMSO controls (upper twopanels), controls treated with cytochalasin D (middle two panels) and in PfACT1 KOs (lower two panels). Seventy-six percent of DMSO control parasitesinvaded erythrocytes in the time frame of the assa. In contrast, 84% of RAP-treated parasites attached to the erythrocyte and could undergo reorientationand appeared to secrete RON proteins which are required for formation of the junction. However, a typical circular junction could never be observed andparasites were incapable of invading erythrocytes demonstrating a critical requirement for parasite actin for host cell invasion.Das S, Lemgruber L, Tay CL, Baum J, Meissner M. Multiple essential functions of Plasmodium falciparum actin-1 during malaria blood-stage development. BMC Biol. 2017 Aug 15;15(1):70.
See original on MMP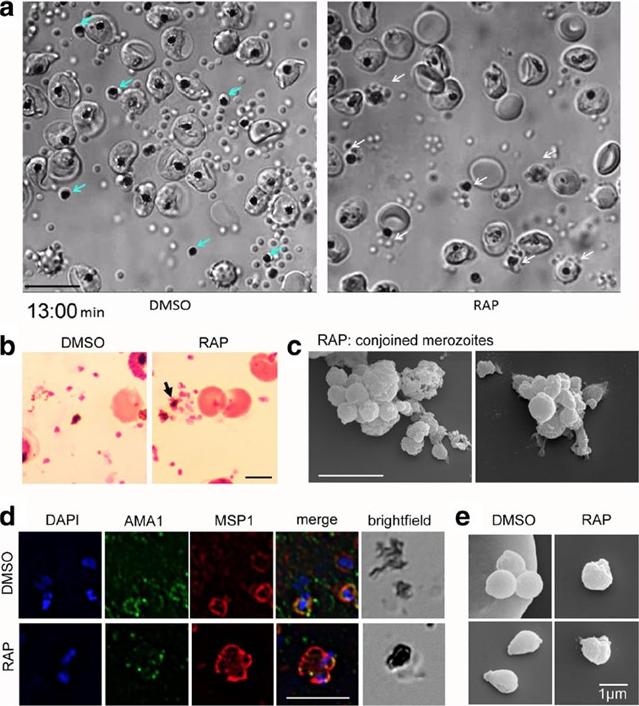
PfACT1 KO merozoites remain conjoined post-egress and possess a dysmorphic ruffled surface. a Still images from Additional file 6: Video S1show normal explosive egress in PfACT1 KOs; however, the RAP-treated population displays conjoined merozoites attached to the FV (white arrows),compared to completely segregated merozoites not attached to the FV (blue arrows) in DMSO controls. b Post-egress, Giemsa-stained RAP-treatedpopulations have parasite structures attached to the FV (black arrow). In DMSO controls, the newly released daughter merozoites are free and notconnected to the FV. Scale bar 5 μm. c Conjoined merozoites with a dysmorphic ruffled surface are apparent by SEM in the RAP-treated population. Scale bar 5 μm. d IFA on post-egress preparations of RAP-treated parasites reveals nuclei (DAPI) and micronemes (AMA1) joined to the FV, with the entire structure bounded by a contiguous plasma membrane (MSP1); defect observed in 76% of all FVs. The FV and merozoites are distinct in DMSO controls (94% of all FVs). N= 21 for DMSO and N = 33 for RAP. Scale bar 5 μm. e Free merozoites are released in the PfACT1 KO population, though they often possess a ruffled surface as observed by SEM. Scale bar 1 μm.Das S, Lemgruber L, Tay CL, Baum J, Meissner M. Multiple essential functions of Plasmodium falciparum actin-1 during malaria blood-stage development. BMC Biol. 2017 Aug 15;15(1):70.
See original on MMP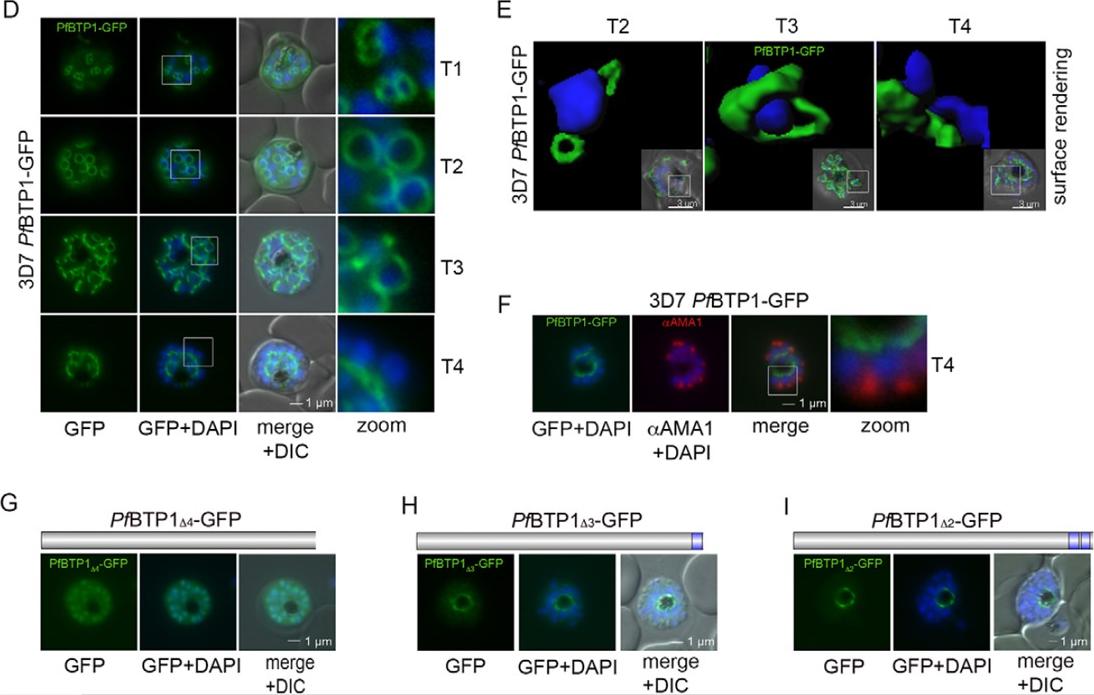
Expression and localization of BTP1. (D) BTP1–GFP can be visualized as a contractile ring structure in unfixed late-stage parasites (T1–T4). It first appears at the apical pole of the nascent merozoite (T1) and ends up at the basal pole of the mature merozoite (T4). Nuclei were stained with DAPI (blue). Enlargement of selected areas are marked with a white square and referred to as ‘zoom’ (a fourfold magnification). (E) Surface-rendering plot of an indicated section, highlighting the spatial relationship between nuclei and the basal-complex ring. (F) Colocalization of BTP1–GFP and the apical membrane antigen 1 (αAMA1; red); AMA1 localizes in the micronomes at the apical pole opposing BPT1, confirming the basal localization of BTP1–GFP at the end of daughter cell formation. (G–I) Localization of the BPT1 deletion mutants. (G) Deletion of all four transmembrane domains leads to a nuclear and cytosolic distribution of BTP1Δ4−GFP. (H) Expression of BTP1 with one (BTP1Δ3−GFP) or two (BTP1Δ2−GFP, I) transmembrane domains results in localization of mutant BTP1 at the food vacuole membrane. DIC, differential interference contrast.Kono M, Heincke D, Wilcke L, Wong TW, Bruns C, Herrmann S, Spielmann T, Gilberger TW. Pellicle formation in the malaria parasite. J Cell Sci. 2016 129(4):673-80.
See original on MMPMore information
| PlasmoDB | PF3D7_1133400 |
| GeneDB | PF3D7_1133400 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | PF11_0344 |
| Orthologs | PBANKA_0915000 , PCHAS_0931000 , PKNH_0931500 , PVP01_0934200 , PVX_092275 , PY17X_0916500 |
| Google Scholar | Search for all mentions of this gene |