PF3D7_1114100 rhomboid protease ROM1 (ROM1)
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. falciparum 3D7 |
Refractory |
USF piggyBac screen (Insert. mut.) | USF PiggyBac Screen | |
| P. berghei ANKA |
Possible |
RMgm-761 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-177 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-176 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
PlasmoGEM (Barseq) | PlasmoGEM | |
| P. yoelii yoelii 17X |
Possible |
RMgm-659 | Imported from RMgmDB | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. berghei ANKA | Asexual |
No difference |
RMgm-761 | Imported from RMgmDB |
| P. berghei ANKA | Asexual |
No difference |
RMgm-177 | Imported from RMgmDB |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-176
Parasitemia develops slower in mice infected with the mutant parasites. Peak parasitemias in mice are comparable to those in wild type infected mice. More then 80% of mice infected with mutant parasites survive infection and clear the parasites from the blood. |
Imported from RMgmDB |
| P. berghei ANKA | Asexual |
No difference |
PlasmoGEM (Barseq) | PlasmoGEM |
| P. yoelii yoelii 17X | Asexual |
Difference from wild-type |
RMgm-659
Analysis of course of blood stage infection in BALB/c mice indicates a mild attenuation of mutant parasites with a decrease in peak parasitemia (25% versus 43%) and a decrease in the duration of infection compared to the wildtype control |
Imported from RMgmDB |
| P. berghei ANKA | Gametocyte |
No difference |
RMgm-761 | Imported from RMgmDB |
| P. berghei ANKA | Gametocyte |
No difference |
RMgm-177 | Imported from RMgmDB |
| P. berghei ANKA | Gametocyte |
No difference |
RMgm-176 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Gametocyte |
No difference |
RMgm-659 | Imported from RMgmDB |
| P. berghei ANKA | Ookinete |
No difference |
RMgm-761 | Imported from RMgmDB |
| P. berghei ANKA | Ookinete |
No difference |
RMgm-177 | Imported from RMgmDB |
| P. berghei ANKA | Ookinete |
Difference from wild-type |
RMgm-176
Normal numbers of ookinetes are produced. A strong reduction in oocyst production. |
Imported from RMgmDB |
| P. berghei ANKA | Oocyst |
No difference |
RMgm-761 | Imported from RMgmDB |
| P. berghei ANKA | Oocyst |
No difference |
RMgm-177 | Imported from RMgmDB |
| P. berghei ANKA | Oocyst |
Difference from wild-type |
RMgm-176
A strong reduction in oocyst production (mean number of oocysts in wild type ranging from 79-213 and in mutants 3-124). Subsequent development of mutant oocysts appears to be normal. The number of mutant sporozoites in oocysts was similar to wild-type oocysts. |
Imported from RMgmDB |
| P. yoelii yoelii 17X | Oocyst |
No difference |
RMgm-659 | Imported from RMgmDB |
| P. berghei ANKA | Sporozoite |
No difference |
RMgm-761 | Imported from RMgmDB |
| P. berghei ANKA | Sporozoite |
Difference from wild-type |
RMgm-176
A strong reduction in oocyst production (mean number of oocysts in wild type ranging from 79-213 and in mutants 3-124). Subsequent development of mutant oocysts appears to be normal. The number of mutant sporozoites in oocysts was similar to wild-type oocysts. No differences of salivary gland invasion of the mutant sporozoites could be detected. Salivary gland sporozoites showed normal gliding motility. The efficiency of liver infection in vivo of the mutant sporozoites was lower (68%) compared to wild type sporozoites. |
Imported from RMgmDB |
| P. berghei ANKA | Sporozoite |
No difference |
RMgm-177 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Sporozoite |
No difference |
RMgm-659 | Imported from RMgmDB |
| P. berghei ANKA | Liver |
Difference from wild-type |
RMgm-761
Liver-stage development was reduced as shown by a 1-day extension of the prepatent period in mice following the inoculation with 10(4) purified sporozoites. While gliding motility and the rate of cell traversal of sporozoites were similar to wild type parasites, in two out of three experiments a reduction in sporozoite in vitro invasion rates was observed that could explain (in part) the delay in the prepatent period. Immunofluorescence analyses of liver stage parasites stained with antibodies against markers for parasite development (HSP70), PVM (UIS4 and EXP1) and merozoite formation (MSP1), revealed no distinct differences in morphology and size between rom1 and wild type liver stages at 24h or 48h after sporozoite invasion. |
Imported from RMgmDB |
| P. berghei ANKA | Liver |
Difference from wild-type |
RMgm-176
The efficiency of liver infection in vivo of the mutant sporozoites was lower (68%) compared to wild type sporozoites. |
Imported from RMgmDB |
| P. berghei ANKA | Liver |
Difference from wild-type |
RMgm-177
Liver-stage development was reduced as shown by a 1-day extension of the prepatent period in mice following the inoculation with 10(4) purified sporozoites.While gliding motility and the rate of cell traversal of sporozoites were similar to wild type parasites, in two out of three experiments a reduction in sporozoite in vitro invasion rates was observed that could explain (in part) the delay in the prepatent period. Immunofluorescence analyses of liver stage parasites stained with antibodies against markers for parasite development (HSP70), PVM (UIS4 and EXP1) and merozoite formation (MSP1), revealed no distinct differences in morphology and size between rom1 and wild type liver stages at 24h or 48h after sporozoite invasion. |
Imported from RMgmDB |
| P. yoelii yoelii 17X | Liver |
Difference from wild-type |
RMgm-659
Sporozoites showed normal motility and traversal and invasion of hepatocytes. However, during development of the liver stages, the number of infected hepatocytes significantly decreased during the first 24 hours compared to wild type liver stages. |
Imported from RMgmDB |
Imaging data (from Malaria Metabolic Pathways)
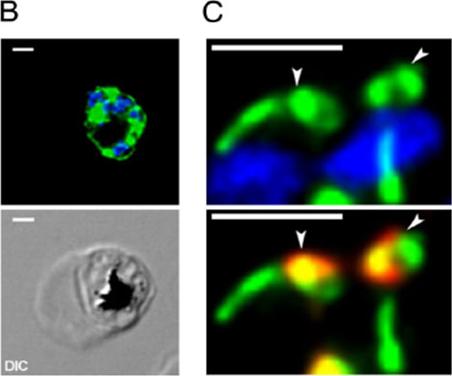
Anti-HA antibodies specifically recognize the HA-PfROM1 protein in transgenic HA-PfROM1 3D7 parasites. (B) IFA analysis of HA-PfROM1 in a developing schizont (4 nuclei stage). (C) Merozoites in a segmenter probed with anti-HA rat mAb 3F10 (green) and the nuclear stain (blue, Upper) or with anti-AMA1 (red, Lower). Note in the individual merozoites anti-HA mAb staining appears to be localized on one side of the nucleus, with characteristic bulbous region (arrowheads) that is toward the apical end of the merozoites. The stalk region runs along one side of the nucleus. PfROM1 was not found associated with micronemes, rhoptries, or dense granules, the three identified secretory organelles of invasion. Release of merozoites from schizonts resulted in the movement of PfROM1 from the lateral asymmetric localization to the merozoite apical pole and the posterior pole. We have named this single thread-like organelle in merozoites, the mononeme.Singh S, Plassmeyer M, Gaur D, Miller LH. Mononeme: a new secretory organelle in Plasmodium falciparum merozoites identified by localization of rhomboid-1 protease. Proc Natl Acad Sci U S A. 2007 104:20043-8
See original on MMP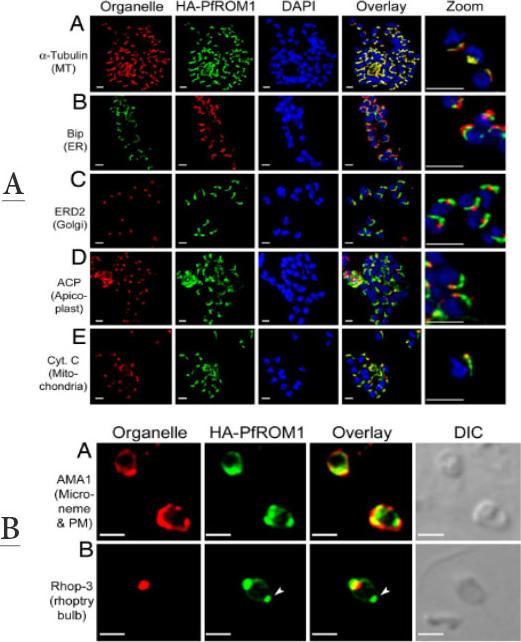
Upper panel: Asymmetric organization of organelles confers a lateral polarity in P. falciparum merozoites. Staining with antibodies against different subcellular organelle markers is shown in the left-hand column. (A) a-Tubulin antibodies staining the merozoite subpellicular microtubules. (B) Bip, an ER marker. (C) ERD2, a cis-Golgi marker. (D) ACP, an apicoplast marker. (E) Cytochrome c, a mitochondrial marker. Corresponding costaining with rat anti-HA mAb 3F10 (A and C–E) or mouse anti-HA mAb 2C16 (B) together with nuclear staining with DAPI (blue) and an overlay of all channels is shown in subsequent columns. The right-hand column shows a digital zoom of merozoites showing that the red and green staining occurs on the same side of the nucleus. (Scale bar, 1 mm.) HA-PfROM1 was observed to be localized in close proximity to longitudinal subpellicular microtubules of the merozoite.Lower panel: HA-PfROM1 is secreted on the merozoite surface and concentrates at the posterior pole of the merozoite. Staining of free merozoites with antibodies against a micronemal marker (A) or with a rhoptry marker at the apical end (B) is shown in the left-hand column (red). Corresponding costaining with rat anti-HA mAb 3F10 (green) together with the overlay between the red and green channels is shown in subsequent columns. The right-hand column shows the corresponding DIC image. Arrow heads point to the merozoite posterior pole. (Scale bar, 1 mm.)Singh S, Plassmeyer M, Gaur D, Miller LH. Mononeme: a new secretory organelle in Plasmodium falciparum merozoites identified by localization of rhomboid-1 protease. Proc Natl Acad Sci U S A. 2007 104:20043-8. Copyright 2009 National Academy of Sciences, U.S.A.
See original on MMPMore information
| PlasmoDB | PF3D7_1114100 |
| GeneDB | PF3D7_1114100 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | PF11_0149, PF11_0150, PF3D7_1114100.1, PF3D7_1114100.2 |
| Orthologs | PBANKA_0933500 , PCHAS_0910800 , PKNH_0911700 , PVP01_0914900 , PVX_091350 , PY17X_0935500 |
| Google Scholar | Search for all mentions of this gene |