PCHAS_1007200 perforin-like protein 1 (PLP1)
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. berghei ANKA |
Possible |
RMgm-1347 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-137 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
RMgm-135 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
PlasmoGEM (Barseq) | PlasmoGEM | |
| P. berghei ANKA |
Possible |
RMgm-4918 | Imported from RMgmDB | |
| P. falciparum 3D7 |
Possible |
28355563 Cell-traversal not possible |
Theo Sanderson, Wellcome Trust Sanger Institute | |
| P. falciparum 3D7 |
Possible |
USF piggyBac screen (Insert. mut.) | USF PiggyBac Screen | |
| P. yoelii yoelii 17X |
Possible |
RMgm-1346 | Imported from RMgmDB | |
| P. yoelii yoelii 17X |
Possible |
RMgm-1348 | Imported from RMgmDB | |
| P. yoelii yoelii 17X |
Possible |
RMgm-4406 | Imported from RMgmDB | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. berghei ANKA | Asexual |
No difference |
RMgm-1347 | Imported from RMgmDB |
| P. berghei ANKA | Asexual |
No difference |
RMgm-137 | Imported from RMgmDB |
| P. berghei ANKA | Asexual |
No difference |
RMgm-135 | Imported from RMgmDB |
| P. berghei ANKA | Asexual |
No difference |
RMgm-4918 | Imported from RMgmDB |
| P. falciparum 3D7 | Asexual |
No difference |
28355563 Cell-traversal not possible |
Theo Sanderson, Wellcome Trust Sanger Institute |
| P. yoelii yoelii 17X | Asexual |
No difference |
RMgm-1346 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Asexual |
No difference |
RMgm-1348 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Asexual |
No difference |
RMgm-4406 | Imported from RMgmDB |
| P. berghei ANKA | Gametocyte |
No difference |
RMgm-1347 | Imported from RMgmDB |
| P. berghei ANKA | Gametocyte |
No difference |
RMgm-137 | Imported from RMgmDB |
| P. berghei ANKA | Gametocyte |
No difference |
RMgm-135 | Imported from RMgmDB |
| P. berghei ANKA | Gametocyte |
No difference |
RMgm-4918 | Imported from RMgmDB |
| P. falciparum 3D7 | Gametocyte |
No difference |
28355563 Cell-traversal not possible |
Theo Sanderson, Wellcome Trust Sanger Institute |
| P. yoelii yoelii 17X | Gametocyte |
No difference |
RMgm-1346 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Gametocyte |
No difference |
RMgm-1348 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Gametocyte |
No difference |
RMgm-4406 | Imported from RMgmDB |
| P. berghei ANKA | Ookinete |
No difference |
RMgm-1347 | Imported from RMgmDB |
| P. berghei ANKA | Ookinete |
No difference |
RMgm-137 | Imported from RMgmDB |
| P. berghei ANKA | Ookinete |
No difference |
RMgm-135 | Imported from RMgmDB |
| P. berghei ANKA | Ookinete |
No difference |
RMgm-4918 | Imported from RMgmDB |
| P. falciparum 3D7 | Ookinete |
No difference |
28355563 Cell-traversal not possible |
Theo Sanderson, Wellcome Trust Sanger Institute |
| P. yoelii yoelii 17X | Ookinete |
No difference |
RMgm-1346 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Ookinete |
No difference |
RMgm-1348 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Ookinete |
No difference |
RMgm-4406 | Imported from RMgmDB |
| P. berghei ANKA | Oocyst |
No difference |
RMgm-1347 | Imported from RMgmDB |
| P. berghei ANKA | Oocyst |
No difference |
RMgm-137 | Imported from RMgmDB |
| P. berghei ANKA | Oocyst |
No difference |
RMgm-135 | Imported from RMgmDB |
| P. berghei ANKA | Oocyst |
No difference |
RMgm-4918 | Imported from RMgmDB |
| P. falciparum 3D7 | Oocyst |
No difference |
28355563 Cell-traversal not possible |
Theo Sanderson, Wellcome Trust Sanger Institute |
| P. yoelii yoelii 17X | Oocyst |
No difference |
RMgm-1348 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Oocyst |
No difference |
RMgm-1346 | Imported from RMgmDB |
| P. yoelii yoelii 17X | Oocyst |
No difference |
RMgm-4406 | Imported from RMgmDB |
| P. berghei ANKA | Sporozoite |
Difference from wild-type |
RMgm-1347
Mutant sporozoites were motile and developed into EEFs in vitro (in Hepa1-6 cells) as efficiently as control parasites, but were poorly infective to mice in vivo, especially when administered through mosquito bites, the natural transmission route. |
Imported from RMgmDB |
| P. berghei ANKA | Sporozoite |
Difference from wild-type |
RMgm-137
Normal numbers of midgut- and salivary gland sporozoites are formed. Sporozoites showed wild type gliding motility. Sporozoites had lost cell passage ability (membrane-damaging capacity) as shown in the cell-wounding assay. Compared to wild type sporozoites, the mutant sporozoites were rapidly immobilized in the dermis. Mutant sporozoites were shown to be arrested by and invade dermal fibroblasts. |
Imported from RMgmDB |
| P. berghei ANKA | Sporozoite |
Difference from wild-type |
RMgm-135
Normal numbers of midgut- and salivary gland sporozoites are formed. Sporozoites have a strongly reduced infectivity to rats after intravenous injection of sprozoites and strongly reduced numbers of exoerythrocytic forms (EEFs) in frozen sections of rat livers were counted 24 h after sporozoite inoculations. |
Imported from RMgmDB |
| P. berghei ANKA | Sporozoite |
Difference from wild-type |
RMgm-4918
Normal numbers of sporozoites; P. falciparum transgene expression in sporozoites; |
Imported from RMgmDB |
| P. falciparum 3D7 | Sporozoite |
Attenuated |
28355563 Cell-traversal not possible |
Theo Sanderson, Wellcome Trust Sanger Institute |
| P. yoelii yoelii 17X | Sporozoite |
Difference from wild-type |
RMgm-1348
Mutant sporozoites were motile and developed into EEFs in vitro (in HepG2/CD81 cells) as efficiently as control parasites, but were poorly infective to mice in vivo, especially when administered through mosquito bites, the natural transmission route.RON4-mCherry is expressed in both oocyst-derived and salivary gland sporozoites, with an apical distribution consistent with rhoptry localization. |
Imported from RMgmDB |
| P. yoelii yoelii 17X | Sporozoite |
Difference from wild-type |
RMgm-1346
Mutant sporozoites were motile and developed into EEFs in vitro (in HepG2/CD81 cells) as efficiently as control parasites, but were poorly infective to mice in vivo, especially when administered through mosquito bites, the natural transmission route. |
Imported from RMgmDB |
| P. yoelii yoelii 17X | Sporozoite |
Difference from wild-type |
RMgm-4406
Normal numbers of salivary gland sporozoites. Injection of 100 sporozoites revealed that Py plp1/spect2 sporozoites were significantly less infectious than Py WT sporozoites (1 out of 10 mice developed blood stage parasitemia). When infection was delivered by five mosquito bites, all Py WT infected mice but none of the Py plp1/spect2 infected mice developed blood stage infection. Increasing the challenge to 15 infectious bites resulted in 7% of Py plp1/spect2 infected mice developing blood stage infection, with a greater than one-day delay to patency observed for infected mice of both groups compared to Py WT. Reduced hepatocyte cell traversal in vitro. Normal/higher hepatocyte invasion rate in vitro compared to wild type. |
Imported from RMgmDB |
| P. berghei ANKA | Liver |
Difference from wild-type |
RMgm-1347
Mutant sporozoites were motile and developed into EEFs in vitro (in Hepa1-6 cells) as efficiently as control parasites, but were poorly infective to mice in vivo, especially when administered through mosquito bites, the natural transmission route. |
Imported from RMgmDB |
| P. berghei ANKA | Liver |
Difference from wild-type |
RMgm-137
No difference was noticed between wild type and mutant parasites in number, size, and fluorescence intensity of EEF at 4, 12, 24, or 48 hr in rat or mouse primary hepatocytes. Sporozoites show in vitro a 'rapid invader' phenotype, as a result of the lack of cell traversal. Mutant sporozoites show an adhesion/attachment phenotype (to CRL-2017 cells in vitro) which is comparable to wild type sporozoites. |
Imported from RMgmDB |
| P. berghei ANKA | Liver |
Difference from wild-type |
RMgm-135
Sporozoites have a strongly reduced infectivity to rats after intravenous injection of sprozoites and strongly reduced numbers of exoerythrocytic forms (EEFs) in frozen sections of rat livers were counted 24 h after sporozoite inoculations. Sporozoites formed normal numbers of EEFs in hepatoma cells (HepG2), indicating that they retain normal infection ability. Sporozoites had completely lost cell passage ability as shown in the cell-wounding assay. Cell-passage activity was estimated by the number of cells wounded by sporozoite passage, which were identified by cytosolic labelling with FITC-conjugated dextran.In Kupffer cell-depleted rats, the sporozoites had restored liver infectivity. Parasitaemia after inoculation of mutant sporozoites was remarkably increased in Kupffer-depleted rats, becoming similar to that after inoculation of wild-type sporozoites In addition, the numbers of EEFs formed in the liver were also similar between spect2-disruptants and the wild type. |
Imported from RMgmDB |
| P. yoelii yoelii 17X | Liver |
Difference from wild-type |
RMgm-1348
Mutant sporozoites were motile and developed into EEFs in vitro (in HepG2/CD81 cells) as efficiently as control parasites, but were poorly infective to mice in vivo, especially when administered through mosquito bites, the natural transmission route.RON4-mCherry is expressed in both oocyst-derived and salivary gland sporozoites, with an apical distribution consistent with rhoptry localization. See additional information about rhoptry discharge and release of RON4-mCherry in transient vacuoles (TVs) and in the PV of infected hepatocytes. |
Imported from RMgmDB |
| P. yoelii yoelii 17X | Liver |
Difference from wild-type |
RMgm-1346
Mutant sporozoites were motile and developed into EEFs in vitro (in HepG2/CD81 cells) as efficiently as control parasites, but were poorly infective to mice in vivo, especially when administered through mosquito bites, the natural transmission route. |
Imported from RMgmDB |
| P. yoelii yoelii 17X | Liver |
Difference from wild-type |
RMgm-4406
Injection of 100 sporozoites revealed that Py plp1/spect2 sporozoites were significantly less infectious than Py WT sporozoites (1 out of 10 mice developed blood stage parasitemia). When infection was delivered by five mosquito bites, all Py WT infected mice but none of the Py plp1/spect2 infected mice developed blood stage infection. Increasing the challenge to 15 infectious bites resulted in 7% of Py plp1/spect2 infected mice developing blood stage infection, with a greater than one-day delay to patency observed for infected mice of both groups compared to Py WT. Reduced hepatocyte cell traversal in vitro. Normal/higher hepatocyte invasion rate in vitro compared to wild type. |
Imported from RMgmDB |
Imaging data (from Malaria Metabolic Pathways)
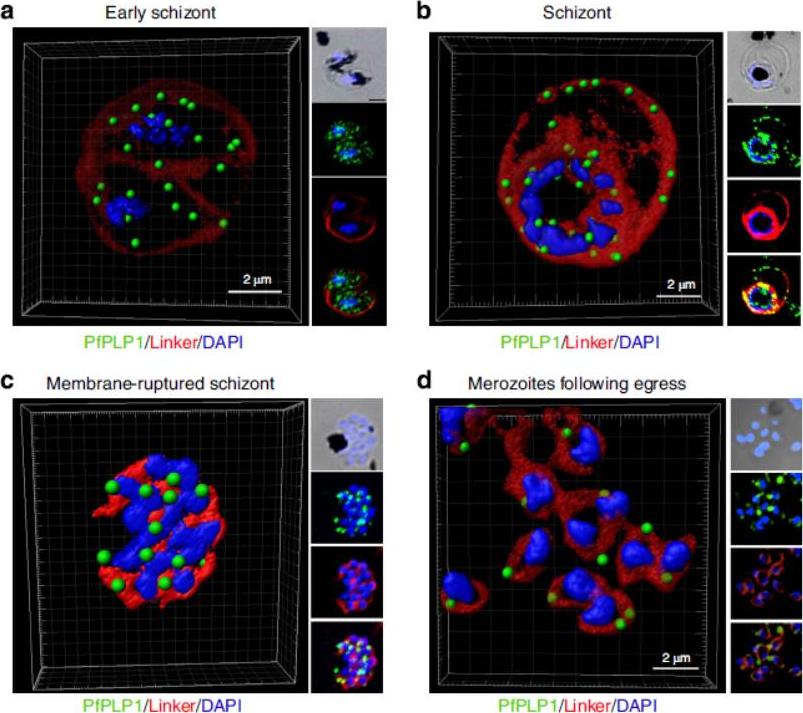
Change in localization of PfPLP1 to host cell membrane during egress of merozoites. (a–d) Schizonts were stained with anti-PfPLP1 followed by anti-mouse Alexa-Fluor 488 (green) and membrane-specific linker dye (red; PKH26 Red Fluorescent Cell Linker Dye) at different time points (B44–48 hpi) during egress. Three-dimensional reconstruction of confocal z-stack images of schizonts was done using Imaris software. PfPLP1 is represented as green dots and membrane dye is shown as two concentric rings corresponding to RBCM and PVM in red. The insets show a bright field and single-slice images with deconvolution. (a) Early schizonts do not show PfPLP1 on the two limiting membranes but shows punctate staining within parasite. (b) Mature schizonts show ring-like staining of PfPLP1 that colocalizes with the membrane dye, suggesting its presence on the membrane. (c) Upon rupture, linker stains the merozoite cell membrane and PfPLP1 shows punctate staining in merozoites with linker staining its cell membrane. (d) After egress, free merozoites show apical localization of PfPLP1. PfPLP1 localizes to RBCM and PVM during egress.Garg S, Agarwal S, Kumar S, Shams Yazdani S, Chitnis CE, Singh S. Calcium-dependent permeabilization of erythrocytes by a perforin-like proteinduring egress of malaria parasites. Nat Commun. 2013 Apr 16;4:1736.
See original on MMP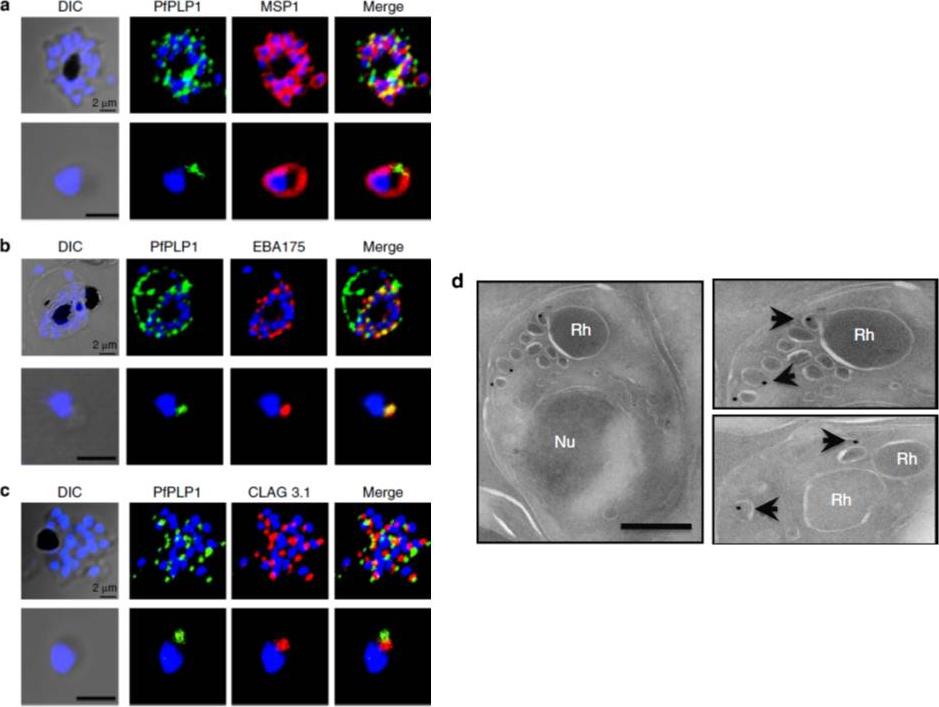
Subcellular localization of PfPLP1 in schizonts and merozoites of P. falciparum. (a) Co-immunostaining of PfPLP1 (green) with merozoite surface protein MSP1 (red). (b) Co-immunostaining of PfPLP1 (green) with micronemal protein EBA-175 (red). PfPLP1 colocalizes with EBA-175 (colocalization coefficient being more than 0.75). (c) Co-immunostaining of PfPLP1 (green) with rhoptry protein CLAG3.1 (red). PfPLP1 is not localized in rhoptry. Nuclei were counterstained with DAPI and Scale bar, 2 mm. (d) Immunoelectron microscopy of PfPLP1. Ultrathin sections of merozoites and late-stage schizonts were labelled with anti-PfPLP1 mouse sera followed by gold-labelled (15 nm) anti-mouse IgG secondary antibodies. Gold particles were detected in elongated, membrane-enclosed micronemes towards the apical end in schizonts and merozoites. Scale bar, 500 nm. PfPLP1 is localized to the micronemes of merozoites.Garg S, Agarwal S, Kumar S, Shams Yazdani S, Chitnis CE, Singh S. Calcium-dependent permeabilization of erythrocytes by a perforin-like protein during egress of malaria parasites. Nat Commun. 2013 Apr 16;4:1736.
See original on MMP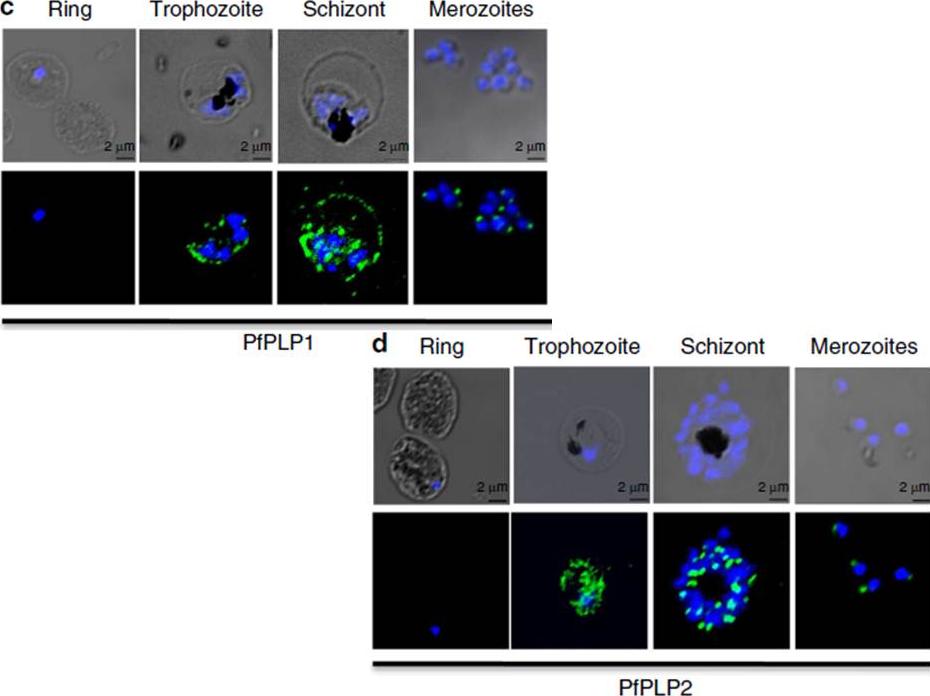
Stage-specific expression of PfPLP1 and PfPLP2 in the blood stage of P. falciparum. Expression of PfPLP1 and PfPLP2 starts in trophozoite stage, with maximal expression in schizonts. In mature schizonts, PfPLP1 and PfPLP2 show punctate staining at the apical end of merozoites. PfPLP1 also shows a ring-like staining, suggesting its localization to RBC membrane. Scale bar, 2 mm.Garg S, Agarwal S, Kumar S, Shams Yazdani S, Chitnis CE, Singh S. Calcium-dependent permeabilization of erythrocytes by a perforin-like protein during egress of malaria parasites. Nat Commun. 2013 Apr 16;4:1736.
See original on MMPMore information
| PlasmoDB | PCHAS_1007200 |
| GeneDB | PCHAS_1007200 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | PC000807.02.0, PC301386.00.0, PC302250.00.0, PCAS_100620, PCHAS_100720 |
| Orthologs | PBANKA_1006300 , PF3D7_0408700 , PKNH_0306700 , PVP01_0308000 , PVX_000810 , PY17X_1007700 |
| Google Scholar | Search for all mentions of this gene |