PKNH_1032300 rhoptry-associated protein 2, putative (RAP2)
Disruptability [+]
| Species | Disruptability | Reference | Submitter | |
|---|---|---|---|---|
| P. berghei ANKA |
Refractory |
RMgm-211 | Imported from RMgmDB | |
| P. berghei ANKA |
Refractory |
RMgm-212 | Imported from RMgmDB | |
| P. berghei ANKA |
Possible |
PlasmoGEM (Barseq) | PlasmoGEM | |
| P. berghei ANKA |
Possible |
RMgm-4428 | Imported from RMgmDB | |
| P. falciparum 3D7 |
Refractory |
USF piggyBac screen (Insert. mut.) | USF PiggyBac Screen | |
Mutant phenotypes [+]
| Species | Stage | Phenotype | Reference | Submitter |
|---|---|---|---|---|
| P. berghei ANKA | Asexual |
Attenuated |
PlasmoGEM (Barseq) | PlasmoGEM |
| P. berghei ANKA | Asexual |
Difference from wild-type |
RMgm-4428
Parasitemia in rap2/3 parasite-infected mice was similar to that in control-parasite-infected mice until day 7 p.i. About 50% of B6 mice infected with rap2/3 parasites showed cerebral complications and died in a similar manner to mice infected with Pb ANKA at days 7-10 p.i., but approximately 40% of the mice ultimately cleared the parasites by day 60 p.i. |
Imported from RMgmDB |
Imaging data (from Malaria Metabolic Pathways)
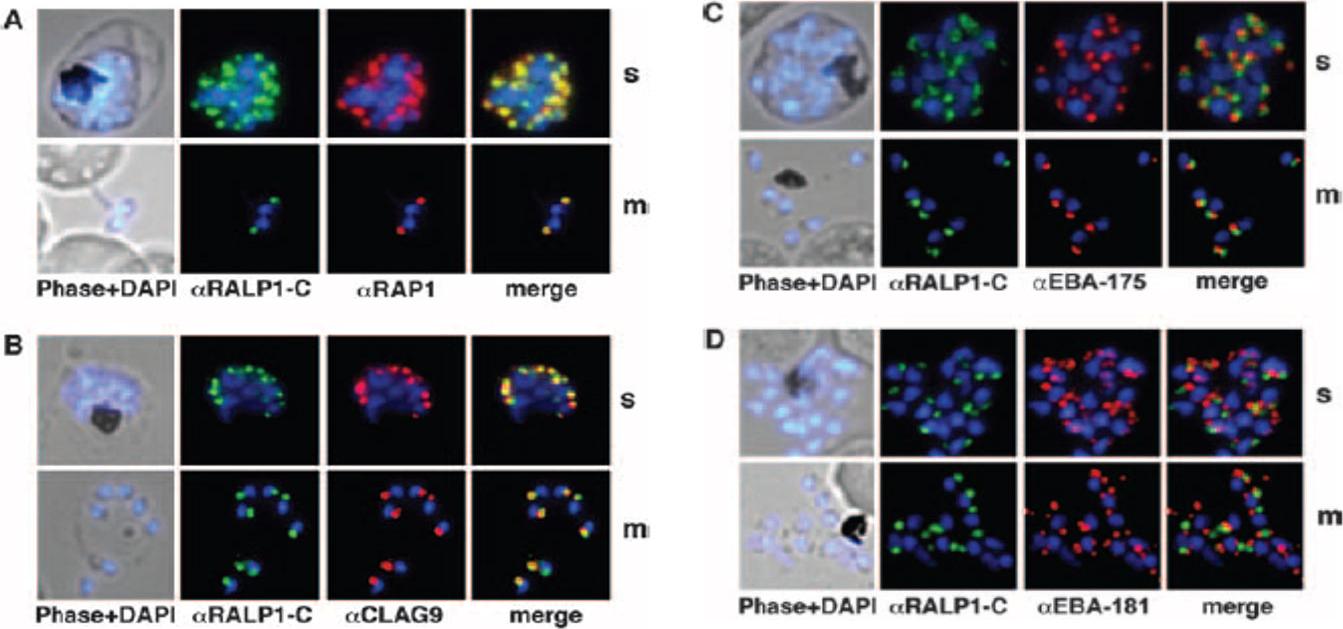
Colocalization studies of RALP1 with rhoptry and microneme marker proteins. (A and B) RALP1-C-specific antibodies (green) colocalize with the rhoptry protein RAP1 (red) (A) and predominantly colocalize with CLAG9 a rhoptry-specific marker, (red) (B) in fixed schizonts (s) and free merozoites (m) using RAP1- and CLAG9-specific antibodies, respectively. (C and D) RALP1-C-specific antibodies (green) visualize a different compartment within the parasite than the microneme marker proteins EBA-175 (red) (C) and EBA-181 (red) (D), as is evident in the merge of the two fluorescence photomicrographs. Nuclei were stained blue (DAPI).Haase S, Cabrera A, Langer C, Treeck M, Struck N, Herrmann S, Jansen PW, Bruchhaus I, Bachmann A, Dias S, Cowman AF, Stunnenberg HG, Spielmann T, Gilberger TW. Characterization of a conserved rhoptry-associated leucine zipper-like protein in the malaria parasite Plasmodium falciparum. Infect Immun. 2008 76:879-87.
See original on MMP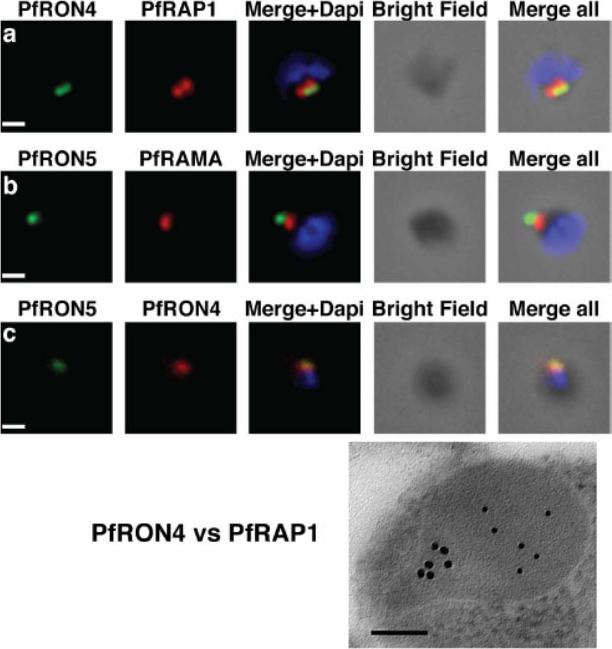
The PfRON4 and PfRON5 proteins localize to the rhoptry neck. A, Antibodies raised against PfRON4 and PfRON5 recognize specific protein products in schizont parasite extracts. B, IFAs using the anti-PfRON antibodies reveal staining of the apical tip of free merozoite, in close apposition with the rhoptry bulb markers PfRAP1 and PfRAMA, suggesting a rhoptry neck localization. Scale bar: 0.2μm C, Immunoelectron microscopy confirms that PfRON 4 localizes to the rhoptry neck. Scale bar: 0.1μmRichard D, Macraild CA, Riglar DT, Chan JA, Foley M, Baum J, Ralph SA, Norton RS, Cowman AF. Interaction between Plasmodium falciparum apical membrane antigen 1 and the Rhoptry neck protein complex defines a key step in the erythrocyte invasion process of malaria parasites. J Biol Chem. 2010 285:14815-22
See original on MMP
MTRAP is a micronemal protein. Upper panel: immunofluorescence microscopy shows apical concentration of MTRAP different to that of PfRAP1, a known basal rhoptry protein. Lower panel, immunoelectron microscopy confirms that apical localization is to the micronemes. m, microneme; R, rhoptry. The arrows indicate concentrations of immunogold labeling against MTRAP-TSR.Baum J, Richard D, Healer J, Rug M, Krnajski Z, Gilberger TW, Green JL, Holder AA, Cowman AF. A conserved molecular motor drives cell invasion and gliding motility across malaria life cycle stages and other apicomplexan parasites. J Biol Chem. 2006 281(8):5197-208.
See original on MMP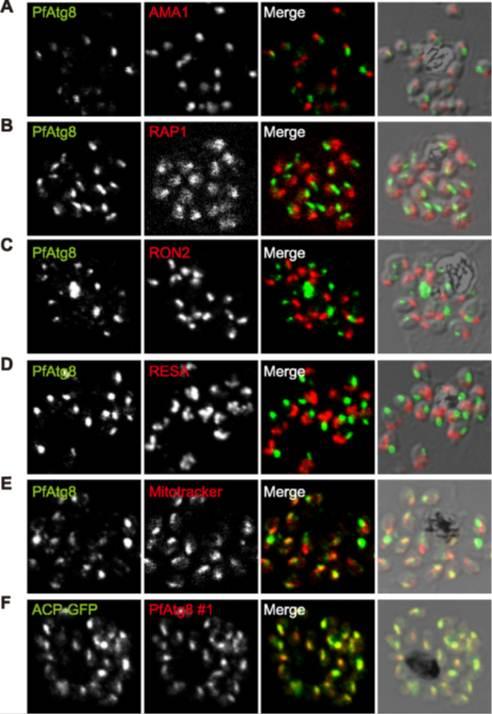
PfAtg8 localizes to the apicoplast. P. falciparum FCR3 (A–E) and P. falciparum 3D7 transfected with ACP-GFP (F–H) were stained with the indicated organelle markers and visualized by confocal microscopy (because ACP-GFP was not uniformly expressed, some merozoites displayed only faint GFP signals). Anti-PfAtg8 antibody #1 was used in (A–F), and anti-PfAtg8 antibody #2 was used in (G). Apical membrane antigen 1 (AMA1) as a microneme marker (A), rhoptry-associated protein 1 (RAP1) as a rhoptry body marker (B), rhoptry neck protein 2 (RON2) as a rhoptry neck marker (C), the ring-infected erythrocyte surface antigen (RESA) as a dense granule marker (D), MitoTrackerRed CMXRos as a mitochondria marker (E), ACPGFP (F–H) and the organellar histone-like protein PfHU (H) as an apicoplast marker were used. Scale bar, 1 mm.Kitamura K, Kishi-Itakura C, Tsuboi T, Sato S, Kita K, Ohta N, Mizushima N. Autophagy-Related Atg8 Localizes to the Apicoplast of the Human Malaria Parasite Plasmodium falciparum. PLoS One. 2012;7(8):e42977
See original on MMP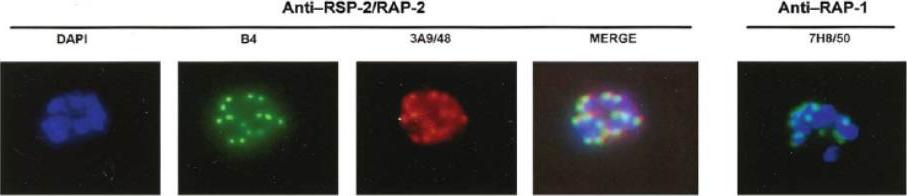
Air-dried immunofluorescence assay (IFA) of D10 schizonts using monoclonal antibodies (MAbs) B4 (anti–ring surface protein [RSP]–2/RAP-2), 3A9/48 (anti–RAP-2), and 7H8/50 (anti RAP-1). The 2 proteins are a part of the rhoptry complex. This complex may either be transferred into the parasitophorous vacuole during the merozoite invasion process (default pathway) or, alternatively, tag erythrocytes during aborted merozoite invasion, which has been observed during in vitro culture.Sterkers Y, Scheidig C, da Rocha M, Lepolard C, Gysin J, Scherf A. Members of the low-molecular-mass rhoptry protein complex of Plasmodium falciparum bind to the surface of normal erythrocytes. J Infect Dis. 2007 196:617-21.
See original on MMP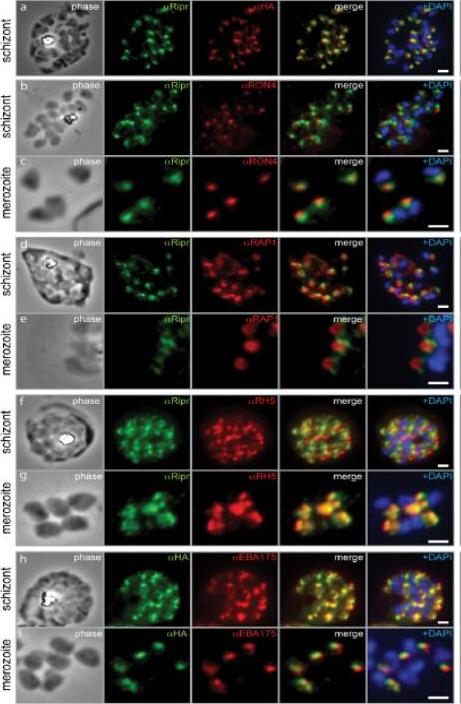
PfRipr localizes to the apical end of merozoites. a) Rabbit polyclonal anti-PfRip antibody (anti-PfRipr/1) recognizes HA-tagged PfRiprHA. b) PfRipr does not co-localize with the rhoptry neck protein RON4 in schizonts. c) PfRipr does not co-localize with the rhoptry neck protein RON4 in merozoites. d) PfRipr does not co-localize with the rhoptry bulb protein, RAP1 in schizonts. e) PfRipr does not co-localize with the rhoptry bulb protein, RAP1 in merozoites. f) PfRipr partially co-localizes with PfRh5 in the schizonts. g) PfRipr mainly co-localizes with PfRh5 in purified merozoites. h) PfRipr co-localizes with the micronemal marker, EBA175, in schizonts. i) PfRipr does not co-localize with the micronemal marker, EBA175, in merozoites.Chen L, Lopaticki S, Riglar DT, Dekiwadia C, Uboldi AD, Tham WH, O'Neill MT, Richard D, Baum J, Ralph SA, Cowman AF. An EGF-like protein forms a complex with PfRh5 and is required for invasion of human erythrocytes by Plasmodium falciparum. PLoS Pathog. 2011 7(9):e1002199.
See original on MMP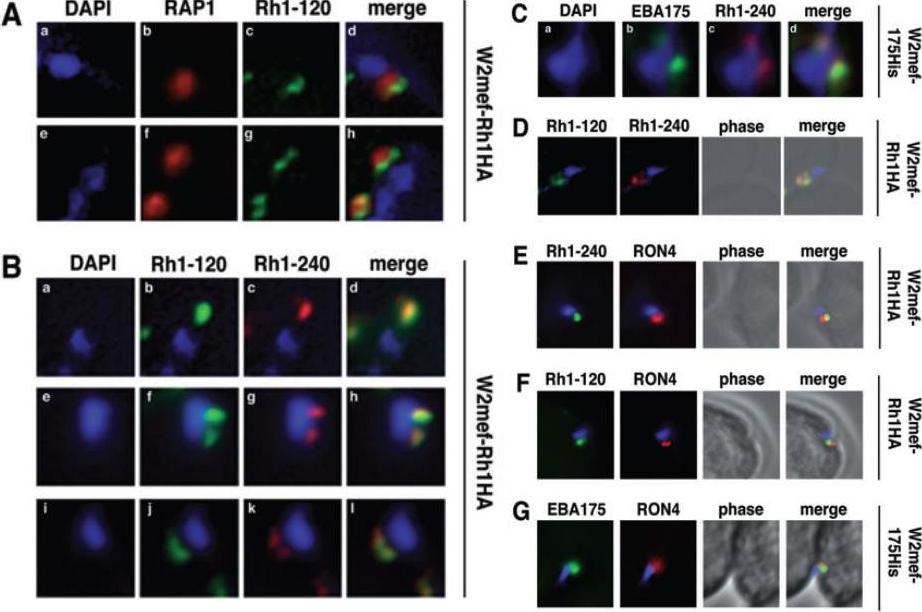
Schizont-processed Rh1-240 colocalizes with Rh1-120 at the apical tip and tight junction of the merozoite. A. The Rh1-120 protein is located at the apical tip and tight junction of merozoites. Free merozoites from the W2mef-Rh1HA line were dual stained with a McAb to RAP1 in the body of the rhoptry and with rat anti-HA antibodies to the tagged 120 kDa Rh1. Anti-HA staining is seen either apical to the RAP1 staining (top panels) or at both sides but posterior to RAP1 staining indicating tight junction staining (bottom panels). Nuclei were stained with DAPI. B. The Rh1-120 protein colocalizes with the Rh1-240 protein at the apical tip and tight junction of merozoites. Free merozoites from the W2mef-Rh1HA line were dual-stained with rat anti-HA antibodies to the tagged 120 kDa Rh1 and with a rabbit antibody (R515) to the 240 kDa Rh1. Colocalization is seen at the apical tip (panels a–d) and at the tight junction (panels e–l) of individual merozoites. W2mef-175His line C. The Rh1-240 protein colocalizes with EBA-175 MAL7P1.176 at the tight junction of merozoites. Free merozoites were dual-stained with mouse anti-His antibodies to tagged EBA-175 and with a rabbit antibody to the 240 kDa Rh1. EBA-175 and Rh1-240 appear to colocalize at the tight junction. D. Free merozoites were dual-stained with rat anti-HA antibodies to the tagged 120 kDa Rh1 and with a rabbit antibody to the 240 kDa Rh1. Simultaneous staining of both the apical tip and the tight junction of a merozoite is seen. E–G. Merozoites were trapped during invasion using cytochalasin D. Rh1-240 was stained with R515, Rh1-120 with rat anti-HA and EBA-175 with mouse anti-His antibodies. PfRON4 was stained either with a mouse antibody (E) or with a rabbit antibody (F and G). Triglia T, Tham WH, Hodder A, Cowman AF. Reticulocyte binding protein homologues are key adhesins during erythrocyte invasion by Plasmodium falciparum. Cell Microbiol. 2009 11:1671-87. Copyright John Wiley & Sons Ltd. 2010.
See original on MMP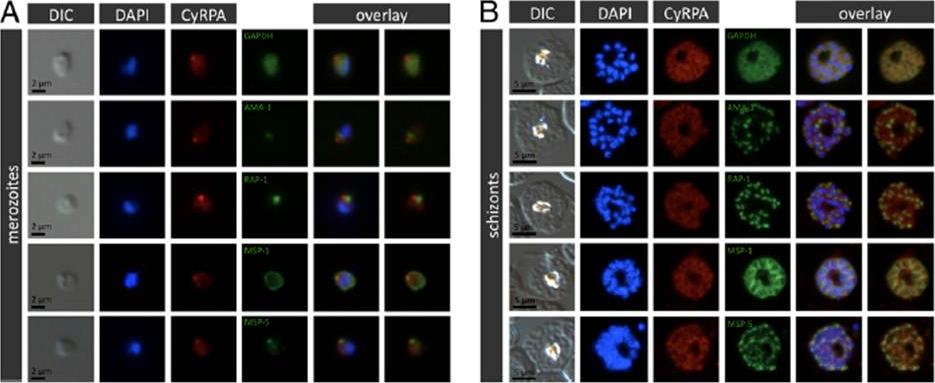
Localization of CyRPA to the merozoite apex by immunofluorescence staining. P. falciparum 3D7 merozoites (A) or schizont stages (B) were coimmunostained with anti-CyRPA mAb c06 (red) and anti-GAPDH Abs (marker for cytosol), AMA-1 (marker for micronemes), RAP-1 (marker for rhoptry bulbs), MSP-1 (marker for merozoite surface), or MSP-5 (green). Nuclei were stained with DAPI (blue). Original magnification x1008. CyRPA is localized at the merozoite apex.Dreyer AM, Matile H, Papastogiannidis P, Kamber J, Favuzza P, Voss TS, Wittlin S, Pluschke G. Passive immunoprotection of Plasmodium falciparum-infected mice designates the CyRPA as candidate malaria vaccine antigen. J Immunol. 2012 188(12):6225-37.
See original on MMP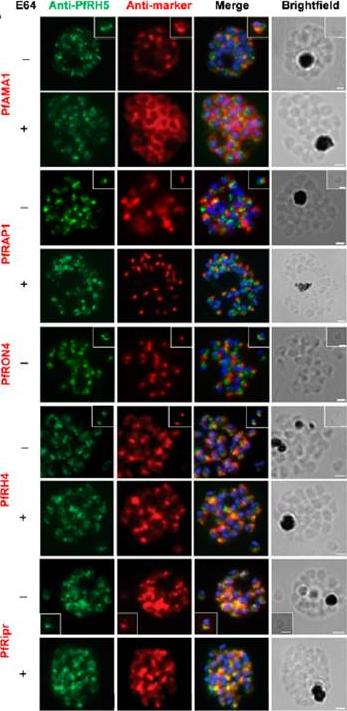
Localization of PfRH5 was assessed by indirect IFA using anti-PfRH5 polyclonal rabbit serum (green). Fixed and permeabilized schizonts with (+) or without (2) E64 treatment or free merozoites (inset) of 3D7 clone P. falciparum were costained with mouse Abs (red) to mark various organelles: PfAMA1 polyclonal (microneme), PfRAP1 mAb (rhoptry body), or PfRON4 mAb (rhoptry neck); or polyclonal mouse serum against further Ags: PfRH4 and PfRipr. Figures show the merge of the dual staining Abs and nuclei stained with DAPI (blue), as well as the brightfield view. Scale bars, 1 mm. anti-PfRH5 rabbit Abs (8) do not colocalize with conventional markers of the rhoptry bulb, rhoptry neck, or micronemes (PfRAP1, PfRON4, and PfAMA1, respectively) in permeabilized schizonts or free merozoites of 3D7 clone P. falciparum parasites. Minimal colocalization in schizonts with PfRH2a/b (data not shown) and also with PfRH4, described as a marker of the rhoptry tip, but significant colocalization with PfRipr in merozoites and late-stage schizonts, especially following treatment with the cysteine protease inhibitor E64, which prevents merozoite release by inhibiting schizont rupture. Impossible to detect PfRH5 on the merozoite surface in any assay, with staining only successful following permeabilization .Douglas AD, Williams AR, Knuepfer E, Illingworth JJ, Furze JM, Crosnier C, Choudhary P, Bustamante LY, Zakutansky SE, Awuah DK, Alanine DG, Theron M, Worth A, Shimkets R, Rayner JC, Holder AA, Wright GJ, Draper SJ. Neutralization of Plasmodium falciparum Merozoites by Antibodies against PfRH5. J Immunol. 2014 192(1):245-58.
See original on MMP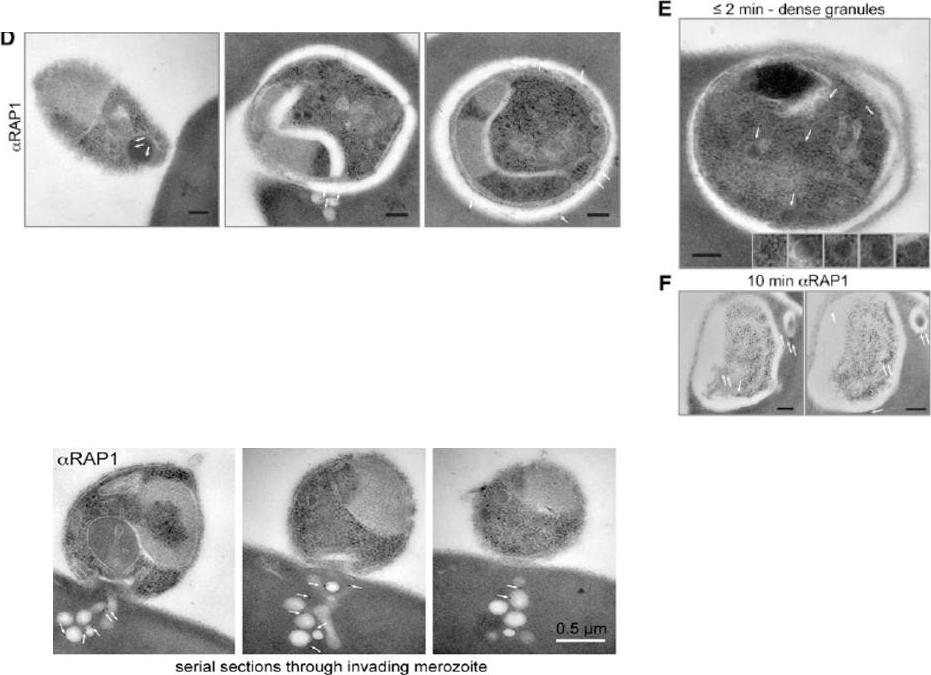
Electron microscopy with immunogold labeling of RAP1 (white arrows). The scale bar represents 0.2 mm. (E) Electron microscopy of merozoite after invasion showing intact dense granules (white arrows and insets). The scale bar represents 0.2 mm. (F) Serial sections of electron microscopy with immunogold labeling of RAP1 10 min after invasion (white arrows). The scale bar represents 0.2 mm. RAP1 labeling revealed prominent extensions of the PV into the erythrocyte cytosol.Lower panel: Serial section electron microscopy with immunogold labeling of RAP1 showing colocalization of RAP1 with rhoptry-derived membrane into erythrocyte cytosol during an untreated merozoite invasion event.Riglar DT, Richard D, Wilson DW, Boyle MJ, Dekiwadia C, Turnbull L, Angrisano F, Marapana DS, Rogers KL, Whitchurch CB, Beeson JG, Cowman AF, Ralph SA, Baum J. Super-resolution dissection of coordinated events during malaria parasite invasion of the human erythrocyte. Cell Host Microbe. 2011 9:9-20.
See original on MMP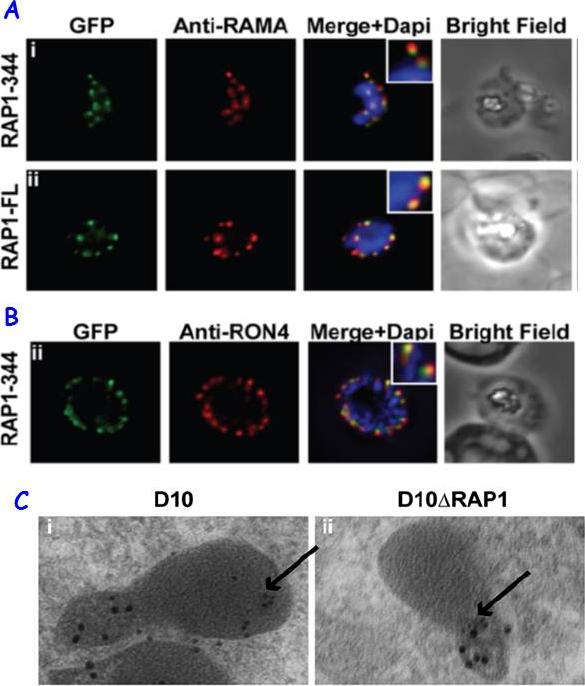
RAP1 contains a bipartite rhoptry signal. A RAP1-344 and RAP1-FL GFP fusions. Both constructs show a punctate fluorescence pattern characteristic of rhoptry localisation. For the RAP1-FL construct, GFP signal overlaps with RAMA. In contrast, the RAP1-344GFP chimera only partially overlaps with RAMA, suggesting rhoptry neck localisation. B. For the RAP1-344 construct, GFP co-localises with the rhoptry neck marker PfRON4. C. Immunoelectron microscopy demonstrates that truncated RAP1 (10 nm beads) in D10DRAP1 parasites is localised in the rhoptry neck, whereas full-length RAP1 in D10 (wild-type) parasites is localised in the rhoptry bulb. PfRON4 PF11_0168 (15 nm beads) is localised in the rhoptry neck in both parasite lines.Richard D, Kats LM, Langer C, Black CG, Mitri K, Boddey JA, Cowman AF, Coppel RL. Identification of Rhoptry Trafficking Determinants and Evidence for a Novel Sorting Mechanism in the Malaria Parasite Plasmodium falciparum. PLoS Pathog. 2009 5(3):e1000328.
See original on MMP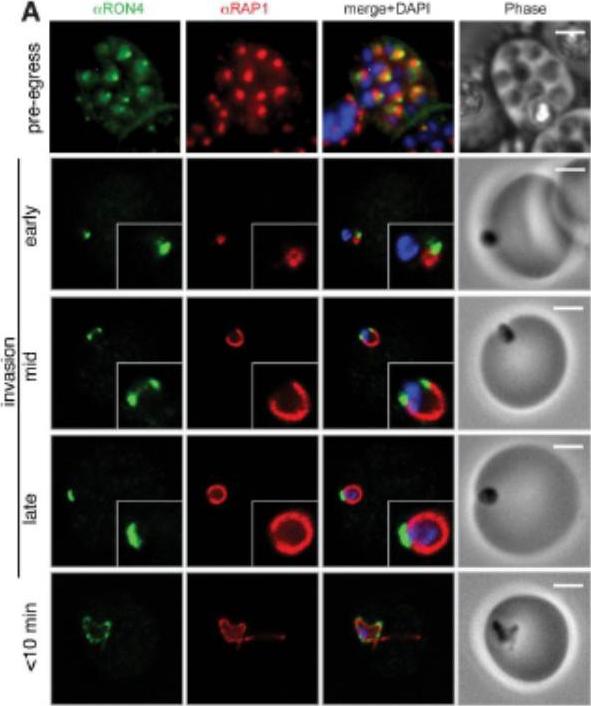
Wide field IFA time course of invasion using A) anti-RAP1/PfRON4. Pre-egress shows standard wide-field image. Invasion images show IFA with deconvolution (single slice). Before invasion commencement, RAP1 clearly remained within the merozoite apex (RAP1 early) posterior to the rhoptry neck as defined by PfRON4. Midinvasion merozoites revealed that RAP1 labeling had moved to an anterior position beginning immediately adjacent to the junction and tracking the exterior of the merozoite as it entered (RAP1 mid). These observations show clearly RAP1 transition pre-, mid-, and postinvasion.Riglar DT, Richard D, Wilson DW, Boyle MJ, Dekiwadia C, Turnbull L, Angrisano F, Marapana DS, Rogers KL, Whitchurch CB, Beeson JG, Cowman AF, Ralph SA, Baum J. Super-resolution dissection of coordinated events during malaria parasite invasion of the human erythrocyte. Cell Host Microbe. 2011 9:9-20.
See original on MMP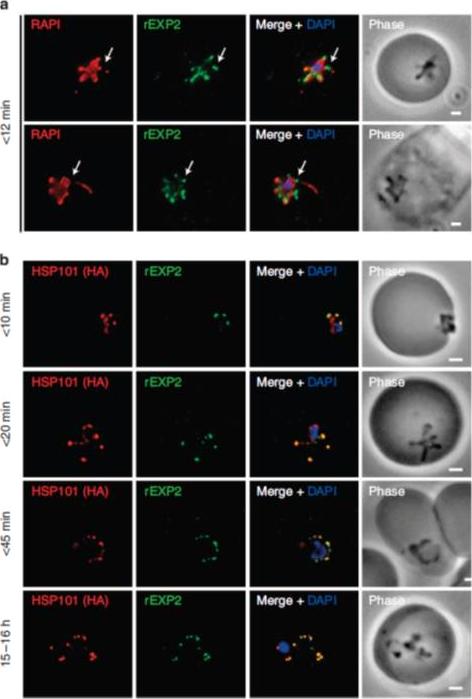
PTEX components localize to the parasite periphery immediately following invasion. (a) Widefield deconvolution imaging of parasites fixed o12 min following erythrocyte invasion and labelled by IFA for EXP2 (green), the PV (RAP1, red) and the nucleus (40,6–diamidino-2-phenylindole (DAPI), blue) showed EXP2 localized in puncta at the parasite periphery at this time point (scale bar, 1 mm). HSP101HA parasites were fixed o10 min, <20 min, <45 min and 15–16 h following erythrocyte invasion and labelled by IFA for EXP2 (green), HSP101HA (HA, red) and the nucleus (DAPI, blue). (b)Widefield deconvolution microscopy (scale bar, 1 mm).Riglar DT, Rogers KL, Hanssen E, Turnbull L, Bullen HE, Charnaud SC, Przyborski J, Gilson PR, Whitchurch CB, Crabb BS, Baum J, Cowman AF. Spatial association with PTEX complexes defines regions for effector export into Plasmodium falciparum-infected erythrocytes. Nat Commun. 2013 Jan 29;4:1415.
See original on MMP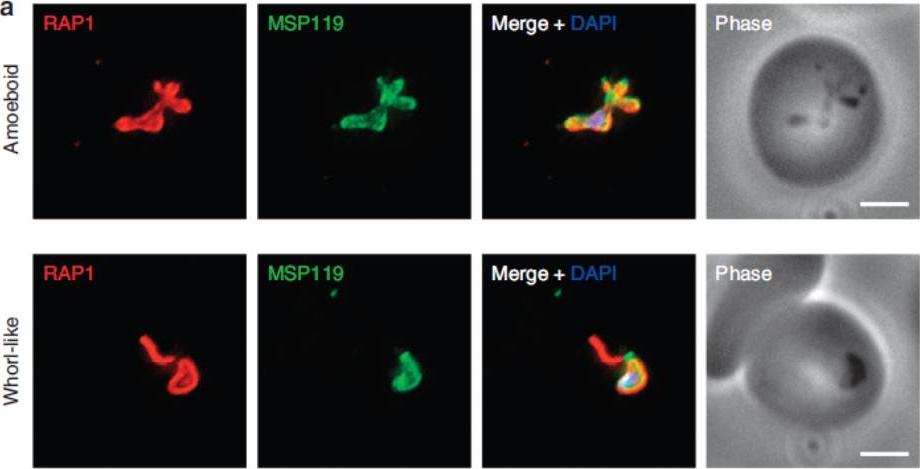
P. falciparum merozoite morphology changes post erythrocyte invasion. Parasites were fixed <12 min following merozoite invasion and labelled by IFA using standard markers for the PV (RAP1, red), parasite plasma membrane (MSP1-19, green) and nucleus (40,6–diamidino-2-phenylindole (DAPI), blue). (a) Widefield deconvolution microscopy identified various morphological changes that had occurred within this period, including pseudopodia-like extensions (top) and whorl-like extensions (bottom). Scale bar, 2.5 mm.Riglar DT, Rogers KL, Hanssen E, Turnbull L, Bullen HE, Charnaud SC, Przyborski J, Gilson PR, Whitchurch CB, Crabb BS, Baum J, Cowman AF. Spatial association with PTEX complexes defines regions for effector export into Plasmodium falciparum-infected erythrocytes. Nat Commun. 2013 Jan 29;4:1415.
See original on MMP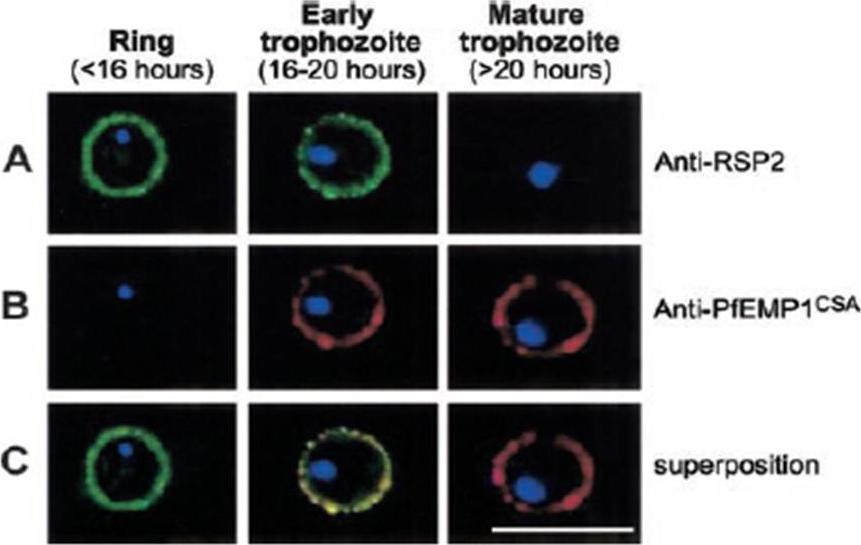
Expression profile of RSP2, the N terminal part of the Rap2 gene, and PfEMP1 at the erythrocyte surface during the blood-stage cycle. RSP-2CSA was stained with mAb B4 (green) and PfEMP1CSA with mAb 1B4/D4 (red) by L-IFA on IE. Parasite nuclei are stained by DAPI (blue). (A) Positive anti–RSP2 staining on synchronized ring stage–infected erythrocytes (16 hours after invasion) and early trophozoites (16 to 20 hours after invasion). No surface staining was detectable on mature forms (> 20 hours after invasion). (B) Absence of PfEMP1 staining on rings (16 hours) but strong IFA signal with early trophozoite and mature stages. (C) Superposition of anti–RSP2 and anti-PfEMP1 staining shows colocalization in early trophozoite stages. Scale bar measures 10 mmDouki JB, Sterkers Y, Lépolard C, Traoré B, Costa FT, Scherf A, Gysin J. Adhesion of normal and Plasmodium falciparum ring-infected erythrocytes to endothelial cells and the placenta involves the rhoptry-derived ring surface protein-2. Blood. 2003 101:5025-32.
See original on MMP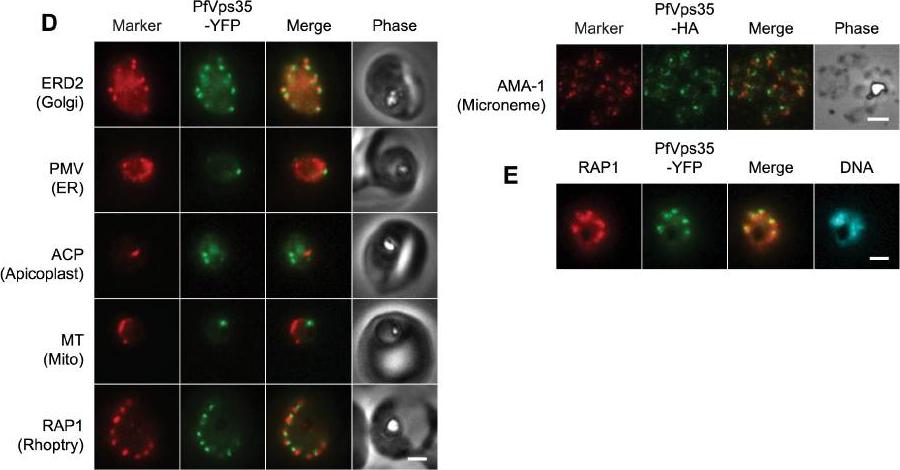
(D) Co-localization of PfVps35-YFP or PfVps35-HA and organellar markers in fixed parasites (except for MitoTracker, which was imaged live). PMV, plasmepsin V; ACP, acyl carrier protein; MT, MitoTracker Red CM-H2Xros; RAP1, rhoptry associated protein 1; AMA1, apical membrane antigen 1. The AMA1 panel shows free merozoites; all others are intraerythrocytic. Organelles labeled by the markers are indicated in parenthesis. Marker-derived fluorescence is pseudocolored red. (E) PfVps35-YFP is adjacent to developing rhoptries in a 2N parasite. Hoechst 33342 fluorescence (DNA) is pseudocolored cyan. In all panels, YFP fluorescence is pseudocolored green. Scale bars, 2 mm.Krai P, Dalal S, Klemba M. Evidence for a Golgi-to-Endosome Protein Sorting Pathway in Plasmodium falciparum. PLoS One. 2014 9(2):e89771.
See original on MMP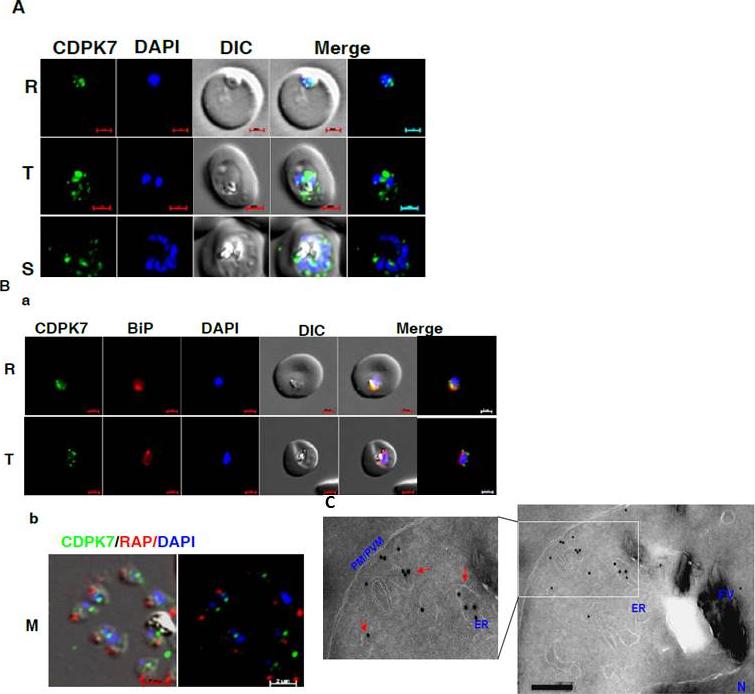
CDPK7 may be present in the ER and vesicles. A IFA was performed to localize CDPK7 in various asexual stages of the parasite using antisera against CDPK7. R, rings; T, trophozoites; S, schizonts. B. Co-staining was done using antisera against CDPK7 and BiP on trophozoites (a) or with anti-RAP1 mAb on free merozoites (b). C. Immuno-EM performed on a trophozoite stage parasite indicated the presence of CDPK7 in ER and vesicular compartments (arrows). Various parasite organelles are indicated: FV, food vacuole; PM/PVM, parasite membrane/parasite vacuolar membrane; ER, Endoplasmic Reticulum; N, Nucleus.Kumar P, Tripathi A, Ranjan R, Halbert J, Gilberger T, Doerig C, Sharma P. Regulation of Plasmodium falciparum development by Calcium-Dependent Protein Kinase 7 (PfCDPK7). J Biol Chem. 2014 Jun 3. [Epub ahead of print]
See original on MMP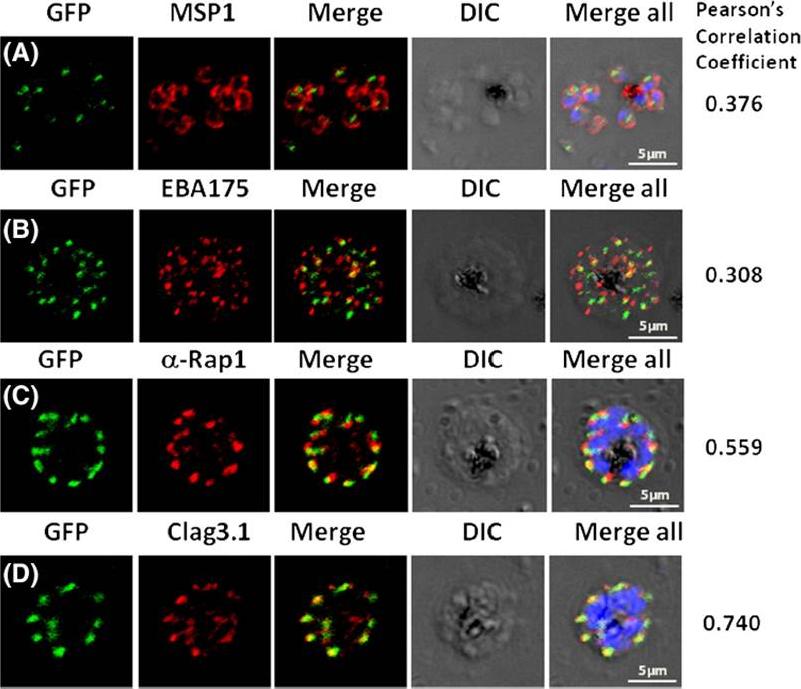
Pfμ1 co-localizeswith rhoptrymarker proteins in schizont stage parasites. Transgenic parasites expressing Pfμ1–GFPwere immunostained with antibodies specific to the Merozoite surface localized MSP1 (A), Microneme localized EBA175 (B), and Rhoptry localized RAP1 (C) and Clag3.1 (D). The parasite nuclei were stained with DAPI and slides were visualized by confocal microscopy. Representative images are shown for each antibody, together with DIC images; scale bars denote 5 μM. To quantify co-localisation, Pearson correlation coefficients of the individual stains were calculated and are shown in the right panel of each image. IFA with antibodies to rhoptry (RAP1 and Clag3.1), microneme (EBA175), and surface markers (MSP1). IFAwith anti-MSP1 antibody showed no overlap in staining betweenMSP1 and Pfμ1 (A). Similar results were seen with antibodies to EBA175 (B). Importantly, anti-RAP1 and anti-Clag3.1 showed co-localization with the Pfμ1–GFP chimeric protein (C and D), suggesting a potential role for Pfμ1 in rhoptry trafficking. Co-localization between Pfμ1 and RAP1 was first observed ~24 h post invasion in budding vesicles near the Golgi. As nuclear division commenced (32 h), Golgi multiplication occurred as well, and this resulted in apical distribution of Pfμ1 along with RAP1 in the rhoptries. Kaderi Kibria KM, Rawat K, Klinger CM, Datta G, Panchal M, Singh S, Iyer GR, Kaur I, Sharma V, Dacks JB, Mohmmed A, Malhotra P. A role for adaptor protein complex 1 in protein targeting to rhoptry organelles in Plasmodium falciparum. Biochim Biophys Acta. 2015 1853(3):699-710.
See original on MMP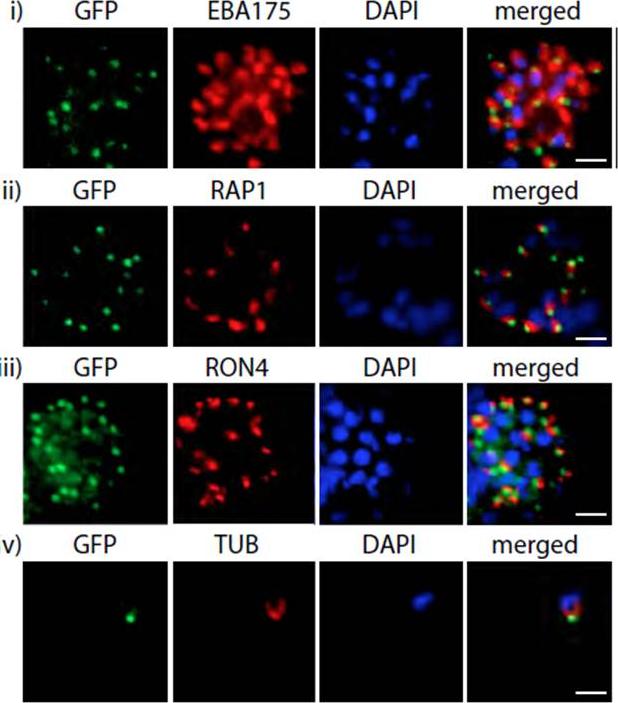
PfMyoB-GFP is located at the apical pole of the merozoite. Indirect immunofluorescence of PfMyoB-GFP (green) with various merozoite organelle markers (red), using antisera to i) EBA175 (microneme marker), ii) RAP1 (rhoptry bulb), iii) RON4 (rhoptry neck) and iv) α-tubulin. Samples were counterstained with DAPI (blue). The merged images are also shown. Rows i-iii show individual schizonts, row iv shows an individual merozoite. Scale bar: 2 μM. Antibodies to GFP produced a compact discrete dot pattern of fluorescence located at the very apical end of the MyoB-GFP parasites, near to the localisation of the apical markers EBA175, RON4, and RAP1. However, whilst in some cases the fluorescent signal partially overlapped with these markers, MyoB appeared to be in a distinct location within the cell, anterior to the microneme marker, the rhoptry bulb, and even to the rhoptry neck.Yusuf NA, Green JL, Wall RJ, Knuepfer E, Moon RW, Schulte-Huxel C, Stanway RR, Martin SR, Howell SA, Douse CH, Cota E, Tate EW, Tewari R, Holder AA. The Plasmodium Class XIV Myosin, MyoB has a Distinct Subcellular Location in Invasive and Motile Stages of the Malaria Parasite, and an Unusual Light Chain. J Biol Chem. 2015 Mar 23.
See original on MMP
Expression and localisation of PfDHHC proteins in P. falciparum schizonts. Triple-HA-tagged PfDHHC proteins were localised by immunofluorescence using antibodies against the 3-HA tag (green). Immunofluorescence staining of each of the tagged PfDHHC proteins was compared against that of the following known localisation markers (red): ERD2 (Golgi marker), BIP (endoplasmic reticulum marker), RAP1 (rhoptry marker), GAP45 (inner membrane complex marker) and MSP1 (plasma membrane marker). Nuclear staining by DAPI is shown in blue. For the staining of PfDHHC9 with GAP45 and MSP1, both a late schizont, as well as an early schizont, is shown in order to differentiate between IMC and plasma membrane localisation. Scale bar: 5 μm.Tay CL, Jones ML, Hodson N, Theron M, Choudhary JS, Rayner JC. Study of Plasmodium falciparum DHHC palmitoyl-transferases identifies a role for PfDHHC9 in gametocytogenesis. Cell Microbiol. 2016 Apr 6.
See original on MMP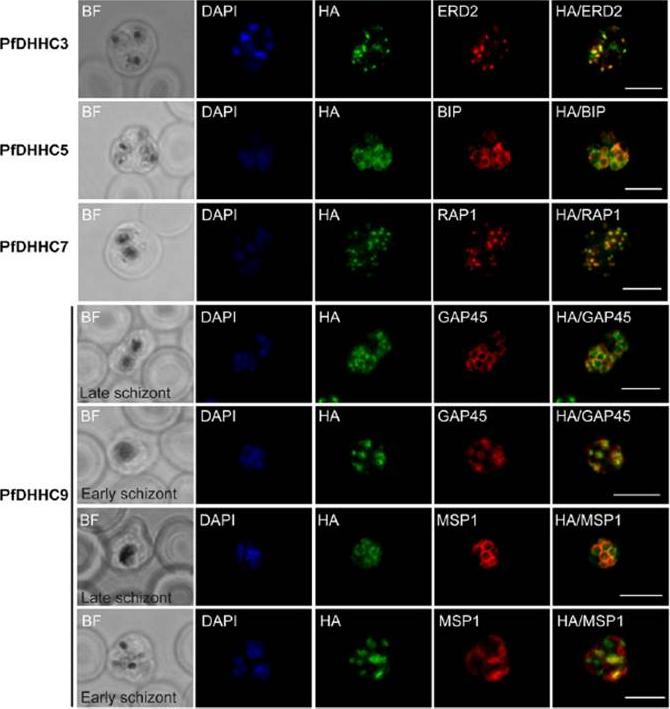
Expression and localization of PfDHHC proteins in Plasmodium falciparum schizonts. Triple-HA-tagged PfDHHC proteins were localized by immuno-fluorescence using antibodies against the 3-HA tag (green). Immunofluorescence staining of each of the tagged PfDHHC proteins was compared against that of the following known localization markers (red): ERD2 (Golgi marker), BIP (endoplasmic reticulum marker), RAP1 (rhoptry marker), GAP45 (inner membrane complex marker) and MSP1 (plasma membrane marker). Nuclear staining by DAPI is shown in blue. For the staining of PfDHHC9 with GAP45 and MSP1, a late schizont, as well as an early schizont, is shown in order to differentiate between inner membrane complex and plasma membrane localization. Scale bar: 5 μm.Tay CL, Jones ML, Hodson N, Theron M, Choudhary JS, Rayner JC. Study of Plasmodium falciparum DHHC palmitoyl transferases identifies a role for PfDHHC9 in gametocytogenesis. Cell Microbiol. 2016 18(11):1596-1610.
See original on MMP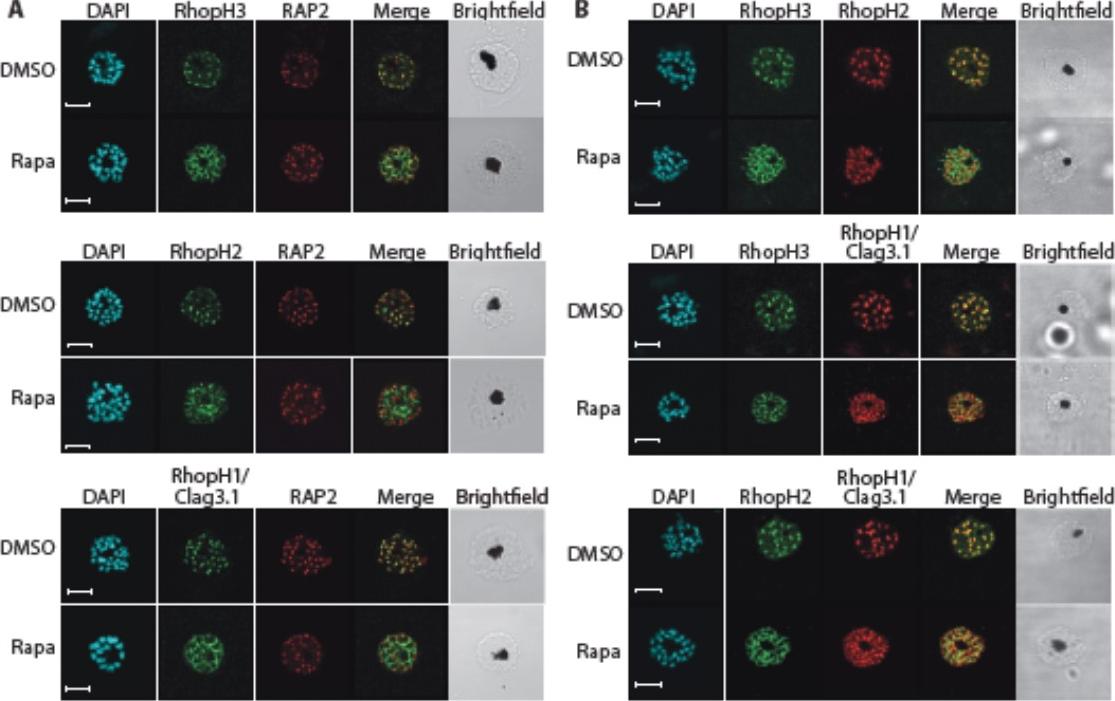
Truncation of RhopH3 leads to mistrafficking of components of the RhopH complex and loss of complex formation. A) IFA showing colocalization of RhopH3, RhopH2 and RhopH1/Clag3.1 with the rhoptry marker RAP2 in schizonts of control (DMSO) rhopH3-loxP parasites but loss of colocalization following rapamycin (Rapa) treatment. Parasite nuclei were visualized by staining with 4,6-diamidino-2-phenylindole (DAPI). Scale bar, 5 μm. Immunofluorescence analysis (IFA) showed that, as expected, RhopH3 colocalized with the rhoptry marker RAP2 in mature schizonts of control rhopH3-loxP parasites. However, in rapamycin-treated (RhopH3Δ4-219 6) parasites, this colocalization was lost, although RAP2 was still detected in a punctate, apically-disposed pattern typical of rhoptries B) Colocalization of the members of the RhopH complex. RhopH3, RhopH2 and Rhop 1/Clag3.1 colocalize in rhopH3-loxP parasites treated with DMSO, but this colocalization is lost in parasites treated with rapamycin. Scale bar, 2 μm. Neither RhopH2 nor RhopH1/Clag3.1 colocalized with 233 RhopH3Δ4-6 in the mutant parasites. The RhopH2 and RhopH1/Clag3.1 signals were also distinct in the mutant parasites, although in this case some limited colocalization of these proteins was apparent (bottom images).Sherling ES, Knuepfer E, Brzostowski JA, Miller LH, Blackman MJ, van Ooij C. The Plasmodium falciparum rhoptry protein RhopH3 plays essential roles in host cell invasion and nutrient uptake. Elife. 2017 6. pii: e23239.
See original on MMPMore information
| PlasmoDB | PKNH_1032300 |
| GeneDB | PKNH_1032300 |
| Malaria Metabolic Pathways | Localisation images Pathways mapped to |
| Previous ID(s) | PK5_0020c, PKH_103190 |
| Orthologs | PBANKA_1101400 , PCHAS_1101100 , PF3D7_0501600 , PVP01_1033400 , PVX_097590 , PY17X_1102500 |
| Google Scholar | Search for all mentions of this gene |